“Despite these brilliant minds, conceptual modeling and empirical research subsequently published by thousands of economists enthused by Nordhaus’ and Romer’s legacies, the global environmental crisis has worsen. The planet’s warming, pollution of the land, air and oceans, and biodiversity loss are ubiquitous in origin. Yet, the markets or innovation technologies have failed to stop the ecocide, or even minimize it. Climate science has been called a hoax and regulations pro nature protection are being ignored or dismantled.”
By Guillermo Paz-y-Miño-C
This year’s Nobel Prize in economics has been awarded to American professors William D. Nordhaus (Yale University) and Paul M. Romer (New York University Stern School of Business) for the integration of “climate change” and “technological innovations” into long-run macroeconomic analyses, respectively.
The Royal Swedish Academy of Sciences, which has granted the “Sveriges Riksbank Prize in Economic Sciences” —the official name— since 1969, highlights that Nordhaus and Romer developed the methods to understand a problem of global relevance: how the economy interacts with nature (exemplified by climate change) and with human knowledge (the ideas and innovations generated to solve problems).
It has long been known to scholars that nature imposes limitations on the economy. At the same time, innovation or “ideas” determine how societies undertake challenges. In the 1990s, Nordhaus introduced the factor “climate” into economic projections. He came up with “DICE,” a Dynamic Integrated Model of Climate and the Economy (watch VIDEO) in which three subcomponents interacted: traditional economic growth theory (markets that produce goods using capital and labor, with natural resources as energy inputs), the carbon cycle (particularly carbon dioxide emissions to the atmosphere derived from burning fossil fuels), and climate (the damage to nature resulting from the accumulation of greenhouse gases).
Separately, and during the 1980s, Romer had observed that technological development correlated with economic prosperity. He asked simple, yet fundamental questions: Where did ideas for new technologies come from? What kind of a product was an idea? Romer proposed that ideas by inventors, engineers or scientists emerged “endogenously” in the marketplace via “rivalry and excludability.” For example, access to inventions like a computer software, a secret soft drink recipe or a coded satellite TV-broadcast could be restricted by encryption (the software or satellite signal) or patent laws (the ownership of the soda formula). For Romer, rivalry and excludability of ideas were central to growth because the latter depended on innovation.
Neither Nordhaus nor Romer offered definitive answers to the challenges of extracting resources from nature with low environmental impact or generating the right amount of knowledge —innovation technologies— to manage such resources to generate sustained and sustainable long-term affluence. In fact, the Committee for the Prize in Economic Sciences noted that the recognition to the researchers was for addressing difficult questions about the economy and providing the conceptual and numerical tools to studying and modeling them.
Nonetheless, based on Nordhaus’ work, corrective measures were suggested to carbon and greenhouse-gases emissions, including carbon taxes on countries. A tactic also rooted in a 1920s notion —in England— that polluters should pay for the damage they caused to society by their polluting practices. A more modern assumption derived from Nordhaus’ research has been that if carbon emissions are limited by law and a high price is set to carbon pollution (by global emissions trading systems), then, minimization of pollution is possible.
Romer’s modeling, on the other hand, later showed that different from the economic growth driven by the accumulation of physical capital (the traditional view), prosperity motivated primarily by the accumulation of ideas did not inevitably experience decreasing returns. He alerted that although unregulated markets will produce technological change, they will tend to underprovide research and development (R&D) and the very goods that R&D could create. To secure global long-run growth, Romer suggested that governments ought to intervene via regulations (patents) and subsidies and incentives to innovation (research). The laws should limit —in time and space— the monopoly rights to goods and balance them with encouragement to creativity.
Despite these brilliant minds, conceptual modeling and empirical research subsequently published by thousands of economists enthused by Nordhaus’ and Romer’s legacies (1980s onwards), the global environmental crisis has worsen (see IPCC October 7, 2018, report). The planet’s warming, pollution of the land, air and oceans, and biodiversity loss are ubiquitous in origin. Yet, the markets or innovation technologies have failed to stop the ecocide, or even minimize it. Climate science has been called a hoax and regulations pro nature protection are being ignored or dismantled in various countries (see reports on the United States A and B).
“Why did New Scientist, ‘the world’s most-read weekly popular science and technology magazine,’ as described on its website, decide to departure from covering ‘international news from a rational, analytical standpoint rooted in the scientific method’ and inject extra doubt into the glyphosate debate? I am referring to the directional ‘probably not’ when swiftly-answering its own query ‘does weedkiller cause cancer?'”
By Guillermo Paz-y-Miño-C
“Does weedkiller cause cancer? Probably not.” These engagement-bite question and answer, as they are known in social media circles when postings lure followers to quick-comment about a topic, were used by New Scientist on Facebook right after a jury in San Francisco concluded that the giant agrochemical and biotech Monsanto must pay $289 million in damages to Dewayne Johnson, who has cancer of the immune system (lymphoma), a condition he and his attorneys claim was caused by exposure to glyphosate-based herbicides commercialized by Monsanto from 1974 to 2000.
The jury’s decision went viral. A 46-year-old school groundskeeper was dying, a world known corporation was being blamed for it, and the state of California offered the perfect stage for litigation. The German Pharmaceutical group Bayer, which back in June, 2018, formalized its engulfing of Monsanto for $60-plus billion, went into frantic damage control, and for a reason. About four thousand other plaintiffs await their day in court. Hundreds of millions, if not billions, of dollars in potential damage awards could be in dispute, more so if Monsanto-Bayer fail in their appeal to the California decision.
But, why did New Scientist, “the world’s most-read weekly popular science and technology magazine,” as described on its website, decide to departure from covering “international news from a rational, analytical standpoint rooted in the scientific method” and inject extra doubt into the glyphosate debate? I am referring to the directional “probably not” when swiftly-answering its own query “does weedkiller cause cancer?”
As the reader might imagine, the New Scientist’s position caused turmoil among academics and science educators, whose mentors and themselves have relied, since 1956, on the London-based enterprise to get their weekly news. For researchers, New Scientist is a classic, like The New York Times or BBC are for journalists.
The concerns in many of the five hundred comments that New Scientist’s Facebook followers wrote were: why did New Scientist appear to align with Monsanto-Bayer, rather than simply apply the scientific method to communicate the facts about glyphosate to the public? Based on the available research, why did New Scientist take the path of “probably not,” rather than an objective “there are some studies suggesting an association between glyphosate and cancer, and others arriving at inconclusive results”? Moreover, why did New Scientist explicitly state in the heading to its post that “there is no evidence that the weedkiller glyphosate causes cancer”? The latter is false; it implies that the studies that have found such indication should be arbitrarily ignored. And that is not how science works.
Some commentators on the New Scientist post added links to the scientific literature and prestigious journals in which associations between glyphosate exposure and cancer had been reported in laboratory animals and limitedly in humans. Others defended Monsanto-Bayer and listed the publications by researchers affiliated with the multi-company Glyphosate Task Force. But the vast majority questioned, not Monsanto-Bayer, but New Scientist for relying on its outreach platform to seed generalized distrust on any probable link between glyphosate and cancer.
The New Scientist’s captions appeared as large texts on a micro-video with images of Mr. Johnson, containers with Round-up (one of the herbicide’s market names), crop fields being labored, activists “impersonating death” and opposing the weedkiller, small airplanes spraying glyphosate, quotes that the International Agency for Research on Cancer (IARC) “considers glyphosate probably carcinogenic” to laboratory animals when exposed in high doses (in direct contradiction to the very New Scientist headings), but that other “studies in humans have found no evidence of a link,” and that “other agencies have concluded there is no increased risk of cancer” due to contact with glyphosate. Yet, the take-home message in the clip was: glyphosate does not or “probably does not” cause cancer (see also companion article “There is no evidence that the weedkiller glyphosate causes cancer…”).
From a rational, analytical standpoint, rigorous scientists would hardly take the “probably not” path. Here is why. It is not a scientific answer. When investigators find evidence, even if limited but of statistical rigor in a controlled study, they state categorically that such evidence exists under the parameters of the research. When no evidence is found, or the numerical sustain is weak, the studies are never declared “probably not” (a “leading-the-mind” hint), but rather inconclusive; and that is Science 101.
“…As for the ‘a’ in the t-shirt, which echoes the pain of a tattoo gone wrong, well, there is not ‘a’ grandeur in this view of life, as per Darwin 1859 (TIES must now produce a clever errata t-shirt amending the misfortune). Neither science is ‘like magic but real,’ as also disseminated by TIES with fervor on Facebook. Nor is the theory of evolution, as presented by Sewell in his misguiding article shared by TIES ‘…a ‘necessary’ truth… not contingent on supporting evidence.’ Nor do ‘Sea Turtles Swim Against the Darwin Current,’ another nonsense from Evolution News that TIES contributed to set in motion in yet another post. — We closed our friendly alert [to TIES] with an ‘Oh My Darwin!!!'”
Typos and errors in scientific publications, or in any long text, are not rare. Even experienced copy editors of journals, magazines and books have their share of faults during volume production. That is why errata exist, to report “wrongs” and, if possible, amend them a posteriori. For example, in our two books, Measuring the Evolution Controversy: A Numerical Analysis of Acceptance of Evolution at America’s Colleges and Universities (2016, best seller 2017), as well as in the recent Kin Recognition in Protists and Other Microbes: Genetics, Evolution, Behavior and Health (2018), we discovered mistakes after publication, even though the publisher and us copy edited and corrected the manuscripts numerous times. We posted the errata online (see Typos and Errors 2016 and 2018) and asked readers to help us spot additional mistakes. Future re-editions will be improved. But keep in mind that our 2016 book was a 198-page and 57,420-word manuscript; and the 2018 volume contained 139,142 words in 428 pages, including +200 figures/sub-figures and tables in each book and their captions (with statistical notation).
“I fully accept the evidence of evolution —including human evolution, but I have to question the grammar on the back of this jacket.”
Although the grammar-correction software available to publishers and authors are powerful enough to detect misspellings, incorrect use of verbs, word redundancy and syntax problems in a text, typos and errors continue to be our most unwanted companions. But errors can be small, sometimes trivial, others substantial, and a few we wish had never been made:
How about introducing error in one of Darwin’s most famous statements “there is grandeur in this view of life” (an eight-word quote from the last paragraph of On The Origin of Species… 1859) and print it on the back of a t-shirt [*] as “Evolution: There is a grandeur in this view of life” (our emphasis on the bold a)? Well, that is precisely what the Teacher Institute for Evolutionary Science (TIES) did, a few weeks ago, in a promotional campaign to “Unlocking the Wonders of Life for Teachers and their Students,” as printed on the front of the t-shirt (in reality, a long-sleeve sweatshirt).
“I fully accept the evidence of evolution —including human evolution, but I have to question the grammar on the back of this jacket” commented one of TIES followers (TP) on Facebook. His wit received likes and smiles [*]. But another (MW) was moved: “Every time I read this I think, such profound words from such a humble man. Makes me shiver every time.”
TIES mission is to “…familiarize interested middle school science teachers with the concepts of natural selection, common ancestry, and diversity in order for them to confidently cover the topics in their classrooms and fulfill their curriculum requirements.” TIES also clarifies that “a middle school science teacher will typically cover many areas of science within his/her annual curriculum, including earth science, physical science, and life science.” And remarks that “it is virtually impossible to become an expert in all of these areas, at least not initially.” Sounds reasonable, however, misquoting Darwin’s ultra famous statement “there is grandeur in this view of life” is a biggie; it denotes cluelessness at best.
“Are we making a big deal out of a silly t-shirt? The ‘a’ in Darwin’s old saying? Below we explain why the ‘a’ symbolizes a pattern of missteps, and there is nothing trivial about them.”
A Google search of Darwin’s phrase gives you 19.5 million hits in 0.28 seconds, at 10:18 AM of a Tuesday in Northeastern United States. In our search, hit number ten corresponded to a 2009 Richard Dawkins‘ video precisely titled “There is grandeur in this view of life,” an impeccable talk delivered at the Atheist Alliance International Conference in Burbank, California.
Are we making a big deal out of a silly t-shirt? The “a” in Darwin’s old saying? After all, it just resembles misquoting Genesis 3 and going to press with “…Let there be light: and there was electricity.” Below we explain why the “a” symbolizes a pattern of missteps, and there is nothing trivial about them.
TIES and Dawkins are connected directly since the Teacher Institute for Evolutionary Science is part of the Center for Inquiry (CFI, a pro secularism organization), which, in turn, is an amalgamation partner of the Richard Dawkins Foundation for Reason & Science (RDFRS). In fact, in 2016, CFI merged with RDFRS. Both organizations originally explained in their websites the rationale (here is the link to F.A.Q. for CFI-RDFRS Merger, but see note below): “…CFI and RDFRS have similar objectives and it makes eminent good sense to combine their resources. CFI’s stated mission is to foster a secular society based on reason, science, and humanist values, and RDFRS shares that goal. And CFI shares the stated mission of RDFRS: to remove the influence of religion in science education and public policy and eliminate the stigma that surrounds atheism and non-belief…” [Note that CFI has a brand new website and this statement from 2016 no longer appears, but in the now-cyber-space-fossil-record CFI had also stated “…By combining their talents, brainpower, and resources, they (CFI-RDFRS) now become the largest freethought organization in the United States. As a result of this merger, they will have greater success in advancing their shared mission. The Richard Dawkins Foundation for Reason & Science will continue as a division of the Center for Inquiry…”].
“Anyone following the evolution wars must have a grasp of the historic players on both sides: evolution versus creationism and its morphs.”
Our discontent with TIES, which has been mounting up for a while (e.g. its ambivalence to openly and up front endorse secularism in science education when interacting with teachers —which is a concern to us, as researchers of the evolution controversy from the perspective of the incompatibility hypothesis and as science educators), reached lava-flow level this past Memorial-Day weekend after TIES posted on its Facebook page a link to a pseudo-science and pseudo-philosophy article by Granville Sewell “Why Evolution is More Certain than Gravity,” an attractive yet impostor heading. TIES engaged its Facebook followers with the bait “check this out” and soon the post received +40 likes and 12 shares [*]. Whoever did this at TIES-Facebook had no idea, or forgot, that Evolution News & Science Today, the platform where the Sewell blurb was unleashed, was a news outlet for the Discovery Institute and its Intelligent Design disciples, the writers at Evolution News.
Anyone following the evolution wars must have a grasp of the historic players on both sides: evolution versus creationism and its morphs (design creationism or intelligent design, theistic evolution, creation science, evolutionary creation, young-earth creationism YEC, or BioLogos, all proponents of proximate or ultimate supernatural causation in evolution, or full deniers of evolution, like YEC). And the 2005 Dover-Pennsylvania trial on ID (Tammy Kitzmiller et al. v. Dover Area School District et al.) should be in the memory of those who profess the proper teaching of evolution in America’s classrooms: ID lost in court for violating the rules of science by “invoking and permitting supernatural causation” in matters of evolution, and for “failing to gain acceptance in the scientific community.”
We immediately alerted our Facebook community that TIES had made that mistake (although some educators had already shared the Sewell article without digesting it; we inferred they did it after trusting TIES and assuming that TIES knew what was being disseminated on social media), and copied Bertha Vázquez, TIES Director, on our post (one of us, GPC, did it). We stated that “…we hope she [Bertha] acts on this immediately and instructs her staff to stop making mistakes like this…” We also referred to the “a” in the t-shirt misquoting Darwin as another bout of inattention in TIES’ record (made public weeks earlier when promoting the slogan Unlocking the Wonders…), and added “…if the excuse is that the post [Sewell’s article] just aimed at generating discussion, well there are hundreds of topics available in the news that can be used for the purpose, rather than sharing, without much thought, a ‘check this out’ article written under the umbrella of INTELLIGENT DESIGN, DESIGN CREATIONISM.” We closed our friendly alert with an “Oh My Darwin!!!” [*]. Bertha did not respond, but the TIES’ post was later deleted. Good for TIES and its Director; amending is what science educators ought to do when erring.
“If there is anything that we remember about our first face-to-face exposure to Richard Dawkins, as graduate students back in the 1990s, is that Richard never tolerated brainlessness or sloppiness in science. — We want TIES to succeed, as much as Dawkins’ brave legacy to prevail.”
TIES states in its Facebook “purpose,” that it “…provides busy educators [our emphasis], homeschooling parents, and curious science lovers with an easily accessible online version of our professional development events and other helpful resources…” Hopefully, our observations to TIES and its Director help those in charge to improve their path of action and honor the association with the prominent RDFRS brand, and with Dawkins himself. We want TIES to succeed, as much as Dawkins’ brave legacy to prevail.
If there is anything that we remember —and we remember a lot— about our first face-to-face exposure to Richard Dawkins, as graduate students back in the 1990s, is that Richard never tolerated brainlessness or sloppiness in science. His talks then, as much as now, were a delight, challenging, inspirational and transformative to colleagues and scientists-to-be. And his sharp, unyielding approach to outreaching the public by conveying the plain scientific truth, the power of evidence and nothing else to engage-bait the skeptics of evolution or give them the impression of harmony between reality and faith, influenced our careers —and deeply— as researchers and evolution/science communicators.
All statistics suggest that the American youth is heading toward a more science-based approach to life and living (e.g. Pew Research science and religion; see also Evolution and the Upcoming Challenges of a Predictable Landscape). Thus, TIES must lead the reason and science debate that projects science educators to the future, rather than inaugurate its journey by experimenting with outreach strategies already entertained by the evolution-and-faith accommodationists of the past.
“We wonder why TIES-Facebook is captivated by the writings of the very Dawkins’ adversaries. Is TIES-Facebook aware of how anti-evolution internet memes become viral in social media via blind sharing? BTW, we take for granted that TIES-Facebook knows who coined the term meme.”
As for the “a” in the t-shirt, which echoes the pain of a tattoo gone wrong, well, there is not “a” grandeur in this view of life, as per Darwin 1859 (TIES must now produce a clever errata t-shirt amending the misfortune). Neither science is “like magic but real” (despite its 665 million hits on Google), as also disseminated by TIES with fervor on Facebook (the fact is that science is like science and magic is an illusion). Nor is the theory of evolution, as presented by Sewell in his misguiding article shared by TIES “…a ‘necessary’ truth… not contingent on supporting evidence.” Nor do “Sea Turtles Swim Against the Darwin Current,” another nonsense from Evolution News that TIES contributed to set in motion in yet another post (May 22, 2018), and about which evolutionary biologist and philosopher of science Kirk Fitzhugh commented “You do realize that EvolutionNews is a mouthpiece for the Discovery Institute and intelligent design?” Yet, TIES gave Kirk a like and kept the post; thus, validating it [*]!
But, in hindsight, that is not all. On April 19, 2018, TIES shared [*] “Cambrian Explosion Shrapnel Still Hitting Evolutionary Scenarios” (the article was from March 28, 2018), a potpourri of statements amassed by the Evolution News staff in which the Cambrian proliferation of life forms was mocked via recycling ID’s favorite smoke grenades: the late “bacterial flagellum” (which ID still believes was designed by a Designer as an “irreducibly complex” structure) and the “blind-Darwinian-evolution analogy” twisted —ID-style— to invalidate Dawkins’ 1986 The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe without Design (a fantastic read!). We wonder why TIES-Facebook is captivated by the writings of the very Dawkins’ adversaries. Is TIES-Facebook aware of how anti-evolution internet memes become viral in social media via blind sharing? BTW, we take for granted that TIES-Facebook knows who coined the term meme.
“…Scientific research –not catchphrases– informs us that the evolution controversy in society is a cancerous zombie, difficult to eradicate. And outcomes of the last elections warn us that assaults on science will metastasize in the United States. Not because researchers ‘do not see’ or ‘do see’ a controversy in the science of evolution, or climate change, or the dark fate of our sun. They have no doubt these are realities. But because unrests resurface in communities whenever we have irreconcilable differences around fiction versus facts. And yes, there is a profound conflict between Faith and Science, intrinsic to their fundamental incompatibility…”
It is also true that after a memorable judgment, in 2005, when Intelligent Design lost in court (Dover, Pennsylvania, Kitzmiller et al. versus Dover School District et al. 2005) for violating the rules of science by “invoking and permitting supernatural causation” in matters of evolution, and for “failing to gain acceptance in the scientific community,” the opportunistic idea to “teach the controversy” over evolution in the science class gained track.
The vested interest was to publicize that there was a “scientific disagreement” over evolution (note that this view was a fabrication, since most of today’s scientists take evolution for granted), and that students ought to be exposed to the “dispute” to secure good schooling. By “teaching the controversy,” designers aimed at injecting creationism into the mix.
“…The sinister drive was to keep the Creator-Designer in the classroom. Of course this was –still is– a controversy, political and ideological.”
Thus, the slogan “there is no controversy about evolution” emerged in response to the proposal to “teach the controversy,” which, in turn, insinuated that modern scientists questioned the authenticity of evolution. The sinister drive was to keep the Creator-Designer in the classroom. Of course this was –still is– a controversy, political and ideological (see facts about the debate).
In consequence, the refurbished slogan “there is no controversy” is, for the most part, a wishful tactic to redirect the “rejection of evolution” back at the “rejectors” themselves, under the premise that –in their minds– the “notion of controversy” enhances the very dismissal of evolution. It is a circular, cosmetic reasoning, with little empirical support, but memed in the social media with enthusiasm.
Now, the contemporary academic topic “evolution controversy” (arguably traceable to the Twentieth Century, 1920s onwards) has never been about any sort of “hypothetical rejection” or “acceptance” of evolution by credible researchers (the spectacular majority of them know evolution happens, contrary to Darwin’s generation), rather, it has been –at least in the United States– about the phenomenon of societal dismissal of science/evolution on the grounds of religious beliefs and conservative political ideology. All sound conceptual studies demonstrate this (e.g. A, B, C,D).
The storm has been about, for example, the support to anti-evolution legislation by lawmakers and their constituents; the low acceptance of evolution by the misinformed general public, and by educators at all levels, who teach creationism because they fear embracing proper science (E, F); and, ultimately, the persistence of ignorance despite the extensive access to knowledge. “That” is –it has been as per chronology– “the controversy.”
“…Imagine asserting that there is no societal controversy concerning ‘alternative facts’ versus facts…”
Here is some food for thought. Imagine asserting that there is no –societal– controversy concerning: climate-change deniers and their opposition to data-based projections of extreme weather fluctuations; or about “alternative facts” versus facts; or homeopathic and chiropractic cures versus scientific medicine; or antivaxxers versus proven herd-immunity effects; or faith healing versus surgical oncology; or anti-GMOs versus no-negative-health-effects-scientifically-attributable-to-GMOs; or pray-sex-health/abstinence versus unwanted pregnancies and sexually-transmitted-diseases… just because –allegedly– no serious scientist gives a damn about the “beliefs in non-facts” by those who worship imagination.
In 2014, public educator Bill Nye debated creationist Ken Ham over the legitimacy of evolution. The encounter itself was controversial, it took place at the Creation Museum in Kentucky. Opponents to the match argued that, by debating, Nye would grant notoriety to Ham’s Museum and the Young-Earth-Creationism agenda. The ‘Science Guy’ defeated Ham at the debate, but it is true that private donations poured in to support the exhibits after the televised event. Currently, the Museum struggles financially and faces sharp criticism by the scientific community.
“…evolution, the ‘E’ word… and E-correctness…”
Since the sticky slogan proclaims “there is no controversy,” it is worth asking: are the science education surrogates expected to put aside the “yuge” societal clash between anti-science and science, or between superstition and empirical reality, and equate them to a minor disagreement, a stone in the shoe?
Like evolution, which continues to occur despite our level of understanding or acceptance of it, the societal controversies are factual and must be addressed as such under reason and science principles, not under the hope that, if we concur to post-like-and-share the catchphrase “there is no controversy about _blank_”, we will make the problem –or part of it– go away. And that is the fallacy of slogans coined to go viral regardless of their silliness. Self deception never pays.
“…any ‘acceptor’ of evolution who believes that God was involved –somehow– in the Creation of the universe, or its laws, is a creationist in principle…”
Is the next step to call evolution the “E-word” so that we rarely use it and, therefore, avoid offending someone? To be E-correct so that our students and public love us as educators, at the same time that we smuggle pseudo-science subliminally into their souls via “teaching techniques“? Or, worse, is the companion, accommodating agenda (to the catchphrase) to force-marry Darwin with Faith to secure that believers accept His message?
Creationism and its disciples come in a range of flavors, from Young Earth Biblical Creationists to Design Creationists, and to any position in which the Creator shows up, even vaguely, in the background of causality (i.e. theistic evolution, creation science, evolutionary creation, BioLogos; links above). In other words, any “acceptor” of evolution who believes that God was involved –somehow– in the Creation of the universe, or its laws, is a creationist in principle. And all morphs of creationism are destined to fail because they merge, deceptively, desire with veracity.
“…as for the fortune of the slogan, trash it…”
Scientific research –not mottos– informs us that the evolution controversy in society is a cancerous zombie, difficult to eradicate. And outcomes of the last elections warn us that assaults on science will metastasize in the United States. Not because researchers “do not see” or “do see” a controversy in the science of evolution, or climate change, or the dark fate of our sun. They have no doubt these are realities. But because unrests resurface in communities whenever we have irreconcilable differences around fiction versus facts. And yes, there is a profound conflict between Faith and Science, intrinsic to their fundamental incompatibility.
PHOTO — Clarence Darrow and William Jennings Bryan during the Scopes Trial (i.e. The State of Tennessee v. John Thomas Scopes 1925) in Dayton, Tennessee. At the Scopes “Monkey” Trial, teacher John T. Scopes was accused of violating the TN’s Butler Act, which considered it illegal to teach human evolution in the state’s public schools. The Scopes Trial is iconic in the history of America’s evolution controversy.
“…Colleges and universities must lead freedom of speech, nurture the battlefield of ideas, pursue world-competitive standards, deter grade inflation (a historical, pervasive companion of conflict resolution), stand for science and reason, teach the realities of the cosmos and evolution without spiritual justifications, and be the safe spaces where the trigger warning ‘no ignorance allowed’ is respected…” – GPC
[click on subtitle to be redirected to The Standard Times]
“There’s been a trend around the country of trying to get colleges to disinvite speakers with a different point of view, or disrupt a politician’s rally. Don’t do that, no matter how ridiculous or offensive you might find the things that come out of their mouths. Let them talk. If you don’t, you just make them a victim, and then they can avoid accountability.”
“That doesn’t mean you shouldn’t challenge them. Have the confidence to challenge them, the confidence in the rightness of your position. But listen. Engage. If the other side has a point, learn from them. If they’re wrong, rebut them. Teach them. Beat them on the battlefield of ideas.”
These were President Obama’s remarks at the Howard University commencement ceremony, back in May 2016. He highlighted a nowadays generalized concern in academia: that millennials (the generation born between the 1980s and early 2000s) have grown intolerant of any view contrary to their deepest, yet maturing convictions, a phenomenon documented by Gallup in its latest report “Free Expression On Campus.”
Although “college students believe First Amendment rights remain strong” in the United States, they also hold conflicting views about “shuttering free speech and impeding a free press under certain circumstances.” For instance, even though 81 percent of college students think that freedom of the press is very secure or secure in the country, and 73 percent think likewise about freedom of speech, one in every five college students (22 percent) believe that “to create a positive learning environment for all, it is more important for colleges to prohibit certain speech or expression of viewpoints that are offensive or biased, than to create an open learning environment where students are exposed to all types of speech and viewpoints —even offensive or biased.”
“…boycott actions against free speech may be rooted in capricious interpretations of First Amendment rights by students active and verbal at their institutions…”
But, what is offensive or biased rhetoric? Slurs and language that intentionally seek to hurt or offend (69 percent of students think so), or the wearing of costumes that stereotype racial or ethnic groups (63 percent believe that), or political views that may upset or offend (27 percent of students think colleges should limit such speech). The Gallup data, therefore, implies that boycott actions against free speech may be rooted in capricious interpretations of First Amendment rights by students active and verbal at their institutions.
Should the media cover protests held at colleges and universities? One in every four students (28 percent, mean of men + women) opposes reporters’ coverage of demonstrations in campus. This view is noticeable among women (37 percent) and African Americans (32 percent). Gallup calls this overall occurrence “support of free press rights in the abstract” (or “in principle”) but not in practice. Protesters believe the press will be unfair in its reporting (49 percent say this), they assert to have the right to be left alone (48 percent), and want to tell their own version of the story on the internet and social media (44 percent). In fact, 86 percent of the polled students prefer the social media for allowing them more control over the story.
Why has intolerance of free speech become ubiquitous? Only tentative answers exist to this question. Greg Lukianoff and Jonathan Haidt, authors of “The Coddling of the American Mind,” have taken a psychological path to explain how “in the name of emotional well-being, college students are increasingly demanding protection from words and ideas they don’t like.” The ultimate aim, Lukianoff and Haidt state, “is to turn campuses into ‘safe spaces’ where young adults are shielded from words and ideas that make some uncomfortable. This movement seeks to punish anyone who interferes with that aim, even accidentally.”
“…Emotional reasoning, distorted thinking, magnification of events, and catastrophizing incidents seem to be central to the individual and collective minds of those who perceive offense in daily experiences…”
The “impulse” is called “vindictive protectiveness” and, as described by Lukianoff and Haidt, “it is creating a culture in which everyone must think twice before speaking up, lest they face charges of insensitivity, aggression, or worse.” Emotional reasoning, distorted thinking, magnification of events, and catastrophizing incidents seem to be central to the individual and collective minds of those who perceive offense in daily experiences. Thus, via negative filtering, groups develop a culture which focuses almost exclusively on the unconstructive, and this allows for simpleminded demonization.
“…The challenge is to identify objectively what is intellectually or psychologically damaging and what is mild…”
Universities must be the safe spaces where the trigger warning “no ignorance allowed” is respected – GETTY IMAGES
“The great contribution of ‘Measuring the Evolution Controversy’ is the rich content of data and analysis that asks detailed questions about the social, economic and political backgrounds of those who tend to reject evolution vs. those who accept evolution as science. Paz-y-Miño-C and Espinosa deftly analyze their data drawn from institutions of higher learning in the United States and particularly New England —which stands as a microcosm of the rest of the country, and indeed elsewhere in the world. It is their scientific approach to these issues which makes this book stand out as a uniquely original contribution.” —Niles Eldredge, PhD, Curator Emeritus of Paleontology at The American Museum of Natural History, New York.
“Pro-science activists and educators constantly bemoan the resistance to the teaching of evolution in the United States. All of us have anecdotes about encounters with the public, parents and students who are misinformed by their churches, Religious-Right groups, and creationist organizations. Paz-y-Miño-C and Espinosa present hard data that support the anecdotal evidence. They also show that although anti-evolutionism typically begins with religion, it is a multi-faceted problem that intersects with political and cultural ideologies. Gathered through careful research over a period of years, their data will enable scientists and defenders of science education to comprehend the roots of the evolution controversy and counteract resistance to evolution more strategically and effectively.” — Barbara Forrest, PhD, co-author with Paul R. Gross of Creationism’s Trojan Horse: The Wedge of Intelligent Design (2007), and expert witness for plaintiffs, Kitzmiller et al. v. Dover Area School District (2005).
“The sweet spot of this collection of essays is the interface of science, history and literacy. Paz-y-Miño-C is, in essence, a champion of rationalism and a passionate defender of literacy standards. His essays deftly weave hard survey data and memorable turns of phrase with evocative imagery… While the essays in this collection are vast in coverage —from climate change to energy policy, stem cell research, vaccinations and, especially, evolution— a clear underlying theme emerges: [the author’s] goal is no less than to counter, through the lens of history and the majesty of rationalism, social forces that sanction ignorance, celebrate denial and… continue to diminish our global status in the fields of science and technology.” Jeff Podos, PhD, Professor of Biology, University of Massachusetts Amherst, USA.
“Paz-y-Miño-C is a firm believer in evolutionary processes. He would like to see decisions made on the basis of facts, not unsupported opinion. He abhors and fears irrational thinking, especially ‘the views of those who see evil in truth and menace in the realities discovered by science.’ He marvels at the intricacy and diversity of life, and how it came about through natural selection… and is clearly frustrated by the unwillingness of so many to see the beauty and majesty in this view of the world and all that it explains.” – Jan A. Pechenik, PhD, Professor of Biology, Tufts University, USA, author of The Readable Darwin: The Origin of Species, as Edited for Modern Readers.
Can single-celled organisms distinguish between same (self or clone) and different (diverse clones)? Can they tell apart kin from non-kin? Can they “learn” to discriminate or recognize members of their own versus others’ cellular lineages? Answers to these questions are provided in three articles (first, second, and third) just published (early view) in the Journal of Eukaryotic Microbiology. The studies summarize outcomes of the symposium “Evidence of Taxa-, Clone-, and Kin-discrimination in Protists: Ecological and Evolutionary Implications,” which took place at the VII European Congress of Protistology, organized last summer in partnership with the International Society of Protistologists (ECOP-ISOP 2015), and hosted by the University of Seville, Spain. I participated at this Symposium and co-authored two of the trilogy papers. But my purpose here is to bring attention to the topic “kin discrimination” or “kin recognition” in unicellular organisms, an area of research that has made much progress in the past fifteen years. There are some technicalities in the text below, but I ask the readers to make an effort and try to understand the broad themes, while being patient with the details. Enjoy! – Guillermo Paz-y-Miño-C.
The unicellular eukaryote Entamoeba invadens (a protist) colored with Green or Red markers. When the Green and Red cells fully aggregate in mixed assemblages, they look yellow under the microscope, an indication that the amebas are able to distinguish members of the same clone, without being affected by the coloration. When grown in the laboratory with non-clone members, the amebas only aggregate with same-clone cells, an indication of preference to group with themselves. Photo courtesy of A. Espinosa 2016.
Readers unfamiliar with “protistan” biology might benefit from grasping some terminology (i.e. protist, prokaryote, eukaryote, clone, multicellularity, kin recognition/discrimination), which I explain next by answering simple questions. For those trained in biology, it might be fine to skip the first five subtitles, below, and move on directly to “From multicellular to unicellular: a round trip:”
What is a protist?
Remember that some organisms are made of single cells. Yes, the entire organism is a cell. For example, bacteria (like the E. coli that lives in the human gut) or amebas, which are also microscopic and can be found in a pond. However, one of the main differences between a bacteria and an ameba is that the former has no nucleus, in contrast to the latter that has it. For that reason, bacteria are called prokaryotes (pro = “before” or “prior to” or, in this case, no nucleus) and amebas are called eukaryotes (eu = “well” or “good” or, in this case, with nucleus). However, many other organisms have nucleated cells (and are, therefore, eukaryotes), like a frog, an orchid, a chimpanzee, a maple tree, or a Blue Whale. The term “protist” applies to unicellular eukaryotes, while the amphibian, the plant, or the aquatic mammal are called multicellular eukaryotes, since their bodies are made of billions of nucleated cells.
Now, the term “protist” is tricky because it includes extremely diverse organisms (which earliest ancestors likely emerged two billion years ago), and over the years scientists have realized that many exceptions exist of organisms that, although are not always unicellular in their life cycles, they are (or should be considered) protists. One of them is the social ameba (or amoeba), Dictyostelium, which forms multicellular assemblages (moving “slugs;” watch video) during its life stages, and also lives in nature as a single-celled, free ranging ameba. To learn more about the ambiguities intrinsic to the term “protist,” and for more examples, go here.
What is a clone?
Because unicellular organisms reproduce, in many cases, via simple cell divisions, the resulting progeny is often identical to the parental cell. A single ameba, for example, can give origin to 2 identical daughter cells, which, in turn, can generate 4 cells (i.e. 2, 4, 8… see cartoon on the right). The resulting thousands, or millions, of cells are “clones” (identical copies of each other). Now, during so many cell divisions, mutations that change the sequence of DNA can –and do– occur, making the descendant cells gradually different from the parental ones. Over time, maybe hundreds of years, a cell line could change enough to become a different clone. Moreover, after thousands or millions of years, different species of amebas can emerge, each distinctive from one another and from the species that gave them origin in the past, a phenomenon known as speciation.
What is multicellularity?
Volvox (a green-algae protist) forms large sphere-colonies made of hundreds, or thousands, of individual cells (see daughter colonies inside). Click on image for source.
Perhaps a more didactic question here is ‘where do multicellular organisms (i.e. frogs, orchids, chimps, maple trees, blue whales) come from’? A first, quick answer is that all multicellular organisms known today originated –at some point– in ancient assemblages of unicellular ancestors (traceable to billions of years ago). However, a more cautious answer is that we have a fragmentary understanding about how primitive single-celled eukaryotes took the path (here I mean driven by natural selection) toward permanent associations in immense cellular cooperatives, which we now call multicellular organisms (note that scientists consider the advent of multicellularity a “major evolutionary transition” in the history of Earth). And that is why studying modern protists, like gregarious amebas (in the genus Entamoeba), or facultative social amebas (i.e. not always social, but in response to environmental circumstances), like Dictyostelium, can give us hints about how multicellularity originated. This particular topic is discussed in the three articles published in JEUKMIC (first, second, and third), and to which I refer in this post. But before I get into that, take a look at the image of Volvox (inset). The organism Volvox is a green-algae protist, which forms large sphere-colonies made of hundreds, or thousands, of individual cells. Inside these spheres, daughter colonies develop and, when they mature, the parental spheres bursts and the descendant colonies are released into the aquatic environment, where they continue to grow and proliferate. Today’s multicellular aggregations of protists, like Entamoeba, Dictyostelium or Volvox, give us clues about how multicellularity might have originated in ancient Earth. Moreover, they are good model-systems to study kin recognition or kin discrimination (which include an organism’s skills for grouping and cooperating with the right partners, and behaving altruistically toward them) in the context of the origin and evolution of multicellularity.
What is kin recognition or kin discrimination?
“The ‘field of kin recognition’… has no consensus on definitions or proposed mechanisms, possibly due to the vast diversity of life histories across organisms and their phylogenetic complexities…”
NASA’s twin astronauts Scott, left, and Mark Kelly. Photo: Tony Cenicola. In humans, identical twins are the only natural “clones;” their genetic relatedness is equal to 100% (represented by r = 1.0). However, the rest of us are related to our siblings only by 50%, or r = 0.5. Our relatedness with our parents is the same, r = 0.5. Can the reader tell why? If so, here is a question: what would be your genetic relatedness with an uncle/aunt, or with a grand parent, or with a second cousin?
This topic can be a bit confusing. However, in one of the articles (the first one listed below), the authors explain why: “…The ‘field of kin recognition’… has no consensus on definitions or proposed mechanisms, possibly due to the vast diversity of life histories across organisms and their phylogenetic complexities (here, phylogeny means ancestry, somewhat analogous to genealogy, not of your own family, but rather of distinctive species or kinds of organisms grouped in distinctive categories). [The authors] refer to “recognition” as an organism’s ability to identify kin [family members] versus non-kin [members of another family]; in addition, [the authors] use the term “discrimination” as the capacity to distinguish one clone from another. Because [the authors] discuss instances of taxa- [taxa = in this particular case means species], clone-, and kin-discrimination/recognition in single-celled organisms capable of both discriminating between same and different, and discriminating/recognizing among clones of distinctive [degree of genetic relatedness, like, for example, values of r less than 1.0], [the authors] use these terms together…”
Why is it relevant to study kin recognition or kin discrimination in single-celled organisms?
To answer this question, I will borrow, again, text from the first article: “…Multicellularity is a major evolutionary transition in which single-celled organisms switched from living individually to permanent assemblages. It is possible that multicellularity originated —more than once— in clonality, via a gradual aggregation of closely related cells, capable of recognizing one another by means of chemical cues, and which lived consistently in intimate proximity and benefited from specialized division of labor (i.e. distinctive tissues and organs with given functions). Such specialization included the full allocation of soma-reproduction [soma = the entire body of an organism] to a small population of cells within the soma, the gametes [i.e. ovules, sperm]… Protists are central to the reevaluation of the theoretical framework and concepts in the field of kin recognition, and to research about the origins and evolution of multicellularity...”
From multicellular to unicellular: a roundtrip
The first article (by Paz-y-Miño-C and Espinosa) is a concise review on “Kin Discrimination in Protists: From Many Cells to Single Cells and Backwards.” In it, the authors summarize the current understanding of the genetics of kin discrimination/recognition in unicellular Eukaryotes, and they do it historically by going back in time, to Darwin and his Origin of Species (1859; Darwin speculated* that selection may be applied to the family; kin discrimination/recognition rely on kin-selection theory), and the influential 1960s, when the modern field of kin recognition was, arguably, born. Here is a simplified version of the first article’s abstract:
“During four decades (1960s to 1990s), the conceptualization and experimental design of studies in kin recognition relied on work with multicellular eukaryotes, particularly invertebrates and vertebrates, and some plants. This pioneering research had an animal behavior approach. During the 2000s, work on taxa-, clone- and kin-discrimination and recognition in protists produced genetic and molecular evidence that unicellular organisms could distinguish between same (self or clone) and different (diverse clones), as well as among conspecifics of close or distant genetic relatedness (Table 1, below). Here we discuss some of the research on the genetics of kin discrimination/recognition and highlight the scientific progress made by switching emphasis from investigating multicellular to unicellular systems (and backwards). We document how studies with protists are helping us to understand the microscopic, cellular origins and evolution of the mechanisms of kin discrimination/recognition and their significance for the advent of multicellularity...”
[Click on Table 1, below, to enlarge]
Readers might find the following excerpts from this article quite intriguing, e.g. “learning” in unicellular organisms (is that possible?):
“…In 1899, H. S. Jennings wrote: ‘Paramecium… an animal that learns nothing, that exercises no choice in any respect, that is attracted by nothing and repelled by nothing, that reacts entirely without reference to the position of external objects, that has but one reaction [movement –watch video below] for the most varied stimuli… can hardly be said to have made the first step in the evolution of mind, and we are not compelled to assume consciousness or intelligence in any form to explain its activities.”
Above: movement behavior in Paramecium. This video is 13-min long and shows various types of Paramecium. If you watch it for a couple of minutes, it shall give you an idea about how these ciliateslook like and move.
“Except for mind, consciousness and intelligence, which are not prerequisites for kin discrimination or recognition (both can also operate in a reflex manner: stimulus-response), Jennings was mistaken about his entire characterization of Paramecium. Since the early 1900s, sensitization, trial-and-error learning, and classical or operant conditioning (relevant attributes among some of the multicellular eukaryotes that learn to recognize kin) have been documented in Paramecium; [including] micro-tube-escape swimming behavior via discrimination learning (1910s), habituation to approach baited and un-baited targets using bacteria as food-reinforcer (1950s), and swim-approach behavior toward mild-electrically-charged fields in learning discrimination tasks using positive and punishment reinforcements (2000s).”
And the authors add: “…But, to our knowledge, there is no direct, experimental evidence that protists can rely specifically on sensitization (i.e. the enhancement of a response to an incremental exposure to a stimulus, for example, the differential frequency exposure to kin versus non-kin during a life cycle), trial-and-error learning (i.e. repeated attempts to solve a task until success, for example, attempts to behave altruistically toward kin, and the benefits it entails, versus the costs of maladaptive altruism toward non-kin), or classical or operant conditioning to discriminate between kin and non-kin (i.e. learning to associate a behavioral or chemical cue with the advantages/disadvantages of aggregating, cooperating or reproducing with conspecifics of [diverse degree of relatedness]). All these topics, remain open areas of investigation and experimentation with protists since, like Paramecium, they possess basic sensory perception capabilities, which could have been co-opted [= adapted] during evolution to function in kin discrimination/recognition…”
Social amebas (or facultative social)
The second article (by Strassmann) is a another review, in this case on “Kin Discrimination in Dictyostelium Social Amoebae.” In it, the author recounts her research program on various species of social amebas, including Polysphondylium violaceum, D. purpureum and D. giganteum. Here is a simplified version of the abstract:
“Evolved cooperation is stable only when the benefactor is compensated, either directly or through its relatives. Social amoebae cooperate by forming a mobile multicellular body in which about 20% of participants ultimately dies to form a stalk [watch video below]. This benefits the remaining individuals that become hardy spores at the top of the stalk, together making up [a] fruiting body. In studied species [of social ameba] with stalked migration, P. violaceum, D. purpureum, and D. giganteum, sorting based on clone identity occurs in laboratory mixes, maintaining high relatedness within the fruiting bodies. D. discoideum has unstalked migration where cell fate is not fixed until the slug forms a fruiting body. Laboratory mixes show some degree of both spatial and genotype-based sorting, yet most laboratory fruiting bodies remain chimeric. However, wild fruiting bodies are made up mostly of clonemates. A genetic mechanism for sorting is likely to be cell adhesion genes tgrB1 and tgrC1, which bind to each other. [These genes] are highly variable, as expected for a kin discrimination gene. It is a puzzle that these genes do not cause stronger discrimination between mixed wild clones, but laboratory conditions or strong sorting early in the social stage diminished by later slug fusion could be explanations.”
Above: the amazing videos of social behavior in amebas, by John Bonner, Professor of Biology at Princeton University. He obtained the images as an undergraduate student. This is a 2-min video of historical value, watch it to the end (it turns spectacular).
Schematic phylogeny based on ssrRNA sequences of the Entamoeba clones discussed in the Espinosa et al. article. Free-living: E. moshkovskii Laredo; commensal E. terrapinae and E. dispar; and parasitic E. invadens IP-1, E. invadens VK-1:NS, E. moshkovskii Snake and E. histolytica HM-1:IMSS.
“Studies on clone- and kin-discrimination in protists have proliferated during the past decade. We report clone-recognition experiments in seven Entamoeba [varieties] (E. invadens IP-1, E. invadens VK-1:NS, E. terrapinae, E. moshkovskii Laredo, E. moshkovskii Snake, E. histolytica and E. dispar). First, we characterized morphometrically each clone (length, width, and cell-surface area) and documented how they differed statistically from one another (as per single-variable or canonical-discriminant analyses). Second, we demonstrated that amebas themselves could discriminate self (clone) from different (themselves versus other clones). In mix-cell-line cultures between closely-related (E. invadens IP-1 versus E. invadens VK-1:NS) or distant-phylogenetic clones (E. terrapinae versus E. moshkovskii Laredo), amebas consistently aggregated with same-clone members. Third, we identified six putative cell-signals secreted by the amebas and which known functions in Entamoeba spp. included: cell proliferation, cell adhesion, cell movement, and stress-induced encystation. To our knowledge, this is the first multi-clone characterization of Entamoeba spp. morphometrics, aggregative behavior, and cell-signaling secretion in the context of clone-recognition. Protists allow us to study cell-cell recognition from ecological and evolutionary perspectives. Modern protistan lineages can be central to studies about the origins and evolution of multicellularity.”
Conclusion
Again, I would like to quote the first article: “…because protists are among the most ancient organisms on Earth, belong to multiple taxonomic groups and occupy all environments, they can be central to reexamining traditional hypotheses in the field of kin recognition, reformulating concepts, and generating new knowledge [to our current understanding of the origins and evolution of multicellularity].” – GPC – Evolution Literacy.
You can contact Guillermo Paz-y-Miño-C via email at guillermo.pazyminoc@gmail.com
Protists are among the most ancient organisms on Earth; they belong to multiple taxonomic groups and occupy all environments. Studies with protists can be central to generating new knowledge to our current understanding of the origins and evolution of multicellularity.
* Darwin (1859) speculated about the “puzzle of the sterile social insects,” in which female workers at a nest dedicate their lives to the persistence of the colony (structured around a large progeny), via assisting a fertile queen to reproduce with the available males. Darwin suggested that, in such cases of apparent sacrifice —by the workers— for the good of all, “selection may be applied to the family.” But, in the late 1800s, he could not offer a detailed mechanistic explanation for the latter. Fisher (1930) and Haldane (1932, 1955) wrestled with the genetics and mathematics of altruism and the anecdotic expression “I would lay down my life for two brothers or eight cousins” became legacy of their work. Hamilton (1964) and Maynard-Smith (1964) further reasoned that the ability to discriminate between close and distant genetic relatives could be directly linked to survival and reproductive success, and, ultimately, to kin selection (Maynard-Smith 1964, 1977). — For references, go to source.
If you are on Twitter (or Facebook), you have probably asked yourself two questions: Am I less popular than my social-media friends? Am I less happy than them? These phenomena (i.e. cyber popularity or cyber happiness) are both real and perceptual.
A paper, by Bollen et al. (total four coauthors), which is available as manuscript (The Happiness Paradox: Your Friends Are Happier Than You), caught my attention. The authors provide evidence of complex associations between the “friendship paradox” and the “happiness paradox.” Enjoy. – Guillermo Paz-y-Miño-C
What is the Friendship Paradox? The perception that YOU are less popular than your friends (on average).
What is the Happiness Paradox? The perception that YOU are less happy than your friends (on average).
What is the issue (so what)? The friendship paradox has been documented in the scientific literature (see Suggested Readings below), but the happiness paradox has remained untested, without quantitative demonstration that it actually happens. Or that, if it does, it occurs in association –somehow– with the friendship paradox. The paper by Bollen et al. makes such connection.
To make it easier for our readers, I will summarize the Bollen et al. study in a digestible manner, although the authors themselves do a fine job explaining the theory, the data and the implications of the research to a specialized audience (see PDF of manuscript). But, for the non-scientists, my summary below might help.
The observations in nature (the social environment):
Online social networking is associated with elevated levels of loneliness, anxiety, displeasure, and dissatisfaction. For example, many times you are on Twitter (or Facebook, or alike social media), you experience an overall sensation that your averagecontacts (i.e. friends, people you follow, or your followers) have more friends or followers than you do, or are happier than you are.
When measured in number of friends-connections, most people tend to have fewer friends than their own friends do on average. Why? Watch the short video posted below (1 minute 36 seconds of your time) for a friendly introduction to the Friendship Paradox.
Now, let us examine numerically how the Friendship Paradox emerges from a simple example. Imagine a social network of four people, as depicted in the image below: Ana has 1 friend (Bill). Bill has 3 friends (Ana, Carla and Dan). Carla and Dan have 2 friends each (Bill, and each other).
There are 8 friends in the network (1 + 3 + 2 + 2). Thus, on average, each person has 2 friends (8/4 = 2).
However, how many friends does each person’s friends have (i.e. how many friends of friends)? Ana is connected to +3 friends of friends, via Bill (who has 3 friends), a subtotal of 3 friends of friends. Bill is connect to 1 + 2 + 2 friends of friends (Ana has 1, Carla has 2, and Dan has 2), a sub total of 5 friends of friends. Carla is connected to 3 + 2 friends of friends (Bill has 3 and Dan has 2), a sub total of 5 friends of friends. And Dan is connected to 3 + 2 friends of friends (Bill has 3 and Carla has 2), a sub total of 5 friends of friends. Therefore, the grand total of friends of friends in the network is 18.
What is the average number of friends of friends? We obtain this number by dividing the total number of friends of friends in the network (=18) by the total number of friends in the network (=8), which gives us a value of 2.25. The Friendship Paradox becomes evident for Ana, Carla and Dan since their average number of friends in the network (=2.0) is lower than the average number of friends of friends in the network (=2.25). The phenomenon is particularly acute for Ana since she has a single friend (Bill) and, therefore, her friendship relations are clearly below both the average number of friends in the network (2.0) and the average number of friends of friends in the network (2.25).
This overall sampling bias effect (= the Friendship Paradox) results –in part– from Bill’s presence in the network. Bill has the most friends and is more often counted in the friends of friends tabulation, thus rising the average value (to 2.25).
The hypothesis
At least in part, Bollen et al. tested the hypothesis that the friendship and happiness paradoxes are predictable byproducts (effects) of universal social network connectivity patterns. And that associations between friendship and happiness also derived from such connectivity (note that the Friendship Paradox has been demonstrated numerous times, but Bollen et al. demonstrated it again with their sample to be able to link it to the Happiness Paradox, the central aspect of their paper).
The phenomena “friendship paradox” or “happiness paradox” result from the inherent structural bias in social networks that favor popular individuals (like Bill, above), who are, by definition, more likely to belong to someone’s social circle. For example, when individuals like you and I equate popularity of our cyber-friends with their cyber-prestige, and compare our own popularity to theirs, we tend to increase our levels of dissatisfaction with ourselves (i.e. we tend to enhance, in our minds, the effects of the friendship-paradox- or the happiness-paradox).
The predictions
Bollen et al. predicted that the effects of the friendship paradox will extend beyond popularity. If popular individuals tend to be happier, then their elevated happiness will become more prevalent as well. Thus, friendship-levels and happiness-levels (as per both paradoxes) should correlate, and this relationship should be measurable.
The authors also predicted that unhappiness will contribute more directly to the negative psycho-social effects of social networking, since it will affect how individuals assess their own subjective wellbeing (i.e. general happiness/unhappiness or life satisfaction/dissatisfaction relative to that of others).
The friendship paradox or the happiness paradox will generate predictable distributions of data, as shown in Fig. 1, below:
Figure 1. Predictable distributions of data expected if the Friendship or Happiness paradoxes are supported by the data (redrawn and adapted from Bollen et al.).
The study
Bollen et al. examined 40,000 Twitter users connected by reciprocal friendship (i.e. people that follow each other).
Two distinctive groups were characterized, the Unhappy group and the Happy group, by applying a subjective assessment of their wellbeing (derived from the Twitter-users’ interactions with their cyber-neighbors), which ranged in a scale from -1.0 (unhappy) to +1.0 (happy).
As shown in Fig. 2, below, both the Unhappy and Happy groups experienced evident friendship-paradox effects (i.e. members of both groups did have, or thought that their friends had, more friends, on average, than themselves). Interestingly, the intensity of the friendship paradox was weaker in the Unhappy group (left) than in the Happy group (right).
Figure 2. Friendship-paradox effects on the Unhappy (left) and Happy (right) groups. Note how the intensity of the friendship paradox was weaker in the Unhappy group than in the Happy group. The “clouds” of data correspond to point distributions of each of the 40,000 Twitter users in the study (modified from Bollen et al.).
In addition, as Fig. 3, below, suggests, both the Happy (top) and Unhappy (bottom) groups experienced clear happy-paradox effects. However, the intensity of the happiness paradox was stronger in the Unhappy group (see the very steep slope in the data point distribution, plus the amount of data points over the main line) than in the Happy group (see the not-so-steep slope in the data point distribution, plus the amount of data points over the main line). In essence, the majority of the sampled Twitter users were less happy (or more unhappy) than their friends on average. These findings support the idea that the Happiness Paradox is real.
Figure 3. Scatter point distributions of an individual’s (exemplified by YOU) sense of happiness as function of his/her (exemplified by YOUR) friends’ average sense of happiness (modified from Bollen et al.).
Individual Happiness and average friends’ Happiness were more strongly related within the Unhappy group than within the Happy group (Fig. 3). Bollen et al. refer to the clustering of unhappy users with themselves, and happy users also with themselves, as being homophilic or assortative in their social networking (both are common terms in behavioral sciences).
Figure 4, below, corroborates the finding that Unhappy Twitter users had fewer friends, on average, than the Happy Twitter users had, on average.
Figure 4. Unhappy Twitter users had fewer friends, on average, than the Happy Twitter users had, on average (modified from Bollen et al.).
Figure 5, below, corroborates the finding that the Happiness Paradox affected more the Unhappy Twitter users, on average, than the Happy Twitter users, on average.
Figure 5. The Happiness Paradox affected more the Unhappy Twitter users, on average, than the Happy Twitter users, on average (modified from Bollen et al.).
Paradoxically, however, as shown in Fig. 6, below, there was a weak correlation between individual happiness and individual popularity for the Unhappy group (Pearson’s R = -0.047; range -0.08 to -0.013). For the Happy group, the correlation was more pronounced (Pearson’s R = 0.126; range 0.081 to 0.171).
Figure 6. There was a weak correlation between individual happiness and individual popularity for the Unhappy group. For the Happy group, the correlation was more pronounced (modified from Bollen et al.).
The overall results suggest that unhappy Twitter users were more strongly affected by the lower happiness of their friends, possibly explaining why this group exhibited a stronger Happiness Paradox (Figs. 3 and 5) in the absence of a strong correlation between Happiness and Popularity (Fig. 6).
Observations of decreased happiness among social media users may result directly from a widespread inflated perception of the happiness of one’s friends.
Given the large role that social media (e.g. Twitter, Facebook) plays in the social lives of billions of individuals, the authors suggest that these environments may induce long-term changes in the public’s social behavior and may, over time, alter the very nature of social relations themselves. — EvoLiteracy.
“Darwin Day… signifies the celebration of the achievements of reason, science, and the advancement of human knowledge. The igniting moments in human history when light was brought into our own origins, when understanding that ordinary apes, like Homo, were capable of the extraordinary, of discovering the truth and debunking obscurantism; yet we still struggle to make science the sole guiding star in our survival decisions, the reliable source of concern and joy, the toolkit to plan our departure from Earth –before our Sun in agonizing heat engulfs its nearest orbiting planets– and seek home somewhere else in the cosmos” — Guillermo Paz-y-Miño-C
A few images and Evolution-Literacy links to celebrate International Darwin Day. — An update on our New England Science Public Series Evolution Volumes 1 and 2, including open access to the studies on acceptance of evolution in the United States. — Links to our Incompatibility Hypothesis papers (evolution versus supernatural causation). — A recount of a visit to the Down House (Darwin’s home). — And an article, from 2013, about the “history of Darwin Day” at the US Congress. Enjoy. – GPC
Here are some photos of Darwin’s statue at the British Museum of Natural History in London, taken back in 2010.
Above: an overall view of Darwin’s statue (marble) at the British Museum of Natural History in London.
Above: a close up of Darwin’s rostrum.
Above: and another close up of this beautiful carving on marble.
Above: the main hall at the British Museum of Natural History in London. Darwin’s statue is in the back, just at the end of the first level of the stairs.
Above: the outdoors of the British Museum of Natural History in London.
NOVA Publishers (New York) used one of these images for the cover of our book Evolution Stands Faith Up: Reflections on Evolution’s Wars (2013). “Paz-y-Miño-C doesn’t ask the reader to ‘believe’ in evolution. He provides overwhelming evidence, clearly written, that shows how scientific inquiry leads to important and practical results, while superstition and faith lead nowhere. Although we may not be able to reason someone out of what they were never reasoned into, the author presents a roadmap for those whose minds are open to discover the wonders and beauty of science.” – Herb Silverman, PhD, author of Candidate Without a Prayer: An Autobiography of a Jewish Atheist in the Bible Belt. Find Evolution Stands Faith Up at NOVA: Soft Cover, Barnes & Noble, Amazon.com, Amazon UK.
Update on NESP Series Evolution
The open access New England Science Public Series Evolution continues to be highly downloaded. Here are some updates:
1,670+ downloads of Volume 1, Number 1: Paz-y-Miño-C G & Espinosa A. 2013. Attitudes toward Evolution at New England Colleges and Universities, United States. New England Science Public: Series Evolution 1(1): 1-32 (ISSN: 2326-0971). The authors compile the most significant results of their conceptual and quantitative studies on the patterns of acceptance of evolution at New England colleges and universities, conducted between 2009 and 2012. They examine the views of New England Faculty and Educators of Prospective Teachers (higher-education faculty themselves, specialized in training future teachers) from 35 colleges and universities, as well as a representative sample of College Students from a Public, Private and two Religious institutions who were polled in three areas: the controversy over evolution versus creationism versus Intelligent Design; their understanding of how science and the evolutionary process work; and their personal convictions concerning the evolution and/or creation of humans in the context of their religiosity… Read MORE open access.
430+ downloads of Volume 2, Number 1: Paz-y-Miño-C G & Espinosa A. 2014. Acceptance of Evolution by America’s Educators of Prospective Teachers. New England Science Public: Series Evolution 2(1): 1-92 (ISSN: 2326-0971). In NESP Series Evolution Vol 2 No 1, Paz-y-Miño-C and Espinosa use the conceptual framework of the Incompatibility Hypothesis (i.e. science/evolution and belief in supernatural causation are incompatible) to document the patterns of acceptance of evolution of 495 Educators of Prospective Teachers affiliated with 281 colleges and universities widely distributed in 4 regions, 9 divisions, and 50 states in the United States. These higher-education professionals (65% PhD-, 22% doctorate-holders) were polled in five areas: (i) their views about evolution, creationism and Intelligent Design, (ii) their understanding of how science and the evolutionary process work, (iii) their position about the hypothetical ‘harmony or compatibility’ between science/evolution and supernatural causation, (iv) their awareness of the age of the Earth, its moon, our solar system and the universe, and the application of the concept of evolution to the cosmos, and (v) their personal convictions concerning the evolution and/or creation of humans in the context of the educators’ religiosity… Read MORE open access.
The Incompatibility Hypothesis: evolution vs. supernatural causation
“Like the oil vs. water experiment, evolution and supernatural causation don’t mix. Evolution raises to the surface.”
Supernatural causation (i.e. the belief in a Supreme Being, creator and sustainer of the universe, omnipresent, omnipotent, omniscient) is a cultural pollutant, incompatible with empirical reality. “Belief” disrupts, distorts, delays and/or stops (3Ds+S) the correct comprehension and acceptance of evidence. We have postulated that the controversy over evolution-and-science versus creationism is inherent to the incompatibility between scientific rationalism/empiricism and the belief in supernatural causation. This hypothesis (= incompatibility) helps us understand and explain the everlasting and fluctuating antagonism –in cycles, from moderate to intense opposition during human history– in the relationship between science/evolution and religion… Read MORE and access free PDFs of scientific articles, including Evolution Controversy: A Phenomenon Prompted by the Incompatibility between Science and Religious Beliefs published in the International Journal of Science in Society (abstract below) [PDF].
Evolution Controversy – Science in Society: Paz-y-Miño-C G & Espinosa A. 2015. Evolution Controversy: A Phenomenon Prompted by the Incompatibility between Science and Religious Beliefs. Int. J. Sci. Soc. 7(2). ISSN 1836-6236. The incompatibility between science and the belief in supernatural causation helps us understand why people do not accept evolution. Belief disrupts, distorts, delays, or stops (3Ds + S) the acceptance of scientific evidence. Here we examine the evolution controversy under three predictions of the incompatibility hypothesis. First, chronological-conflict-and-accommodation, which explains the historical re-emergence of antagonism between evolution and religion when advances in science continue to threaten the belief in supernatural causation; in such situations, creationists’ rejection of and subsequent partial acceptance of the new scientific discoveries are expected. Second, change in evolution’s acceptance is a function of educational attainment, which explains the positive association between acceptance of evolution and level of education. And third, change in evolution’s acceptance is a function of religiosity, which explains the negative association between acceptance of evolution and level of religious beliefs… We emphasize that harmonious coexistence between science and religion is illusory. If co-persisting in society, their relationship will fluctuate from moderate to intense antagonism. Read MORE open access [PDF].
At the Down House: Darwin’s Home
“I visited the Down House, Darwin’s Home, in July 2010. Here are a few pictures I wanted to share in celebration of the International Darwin Day, February 12. Prior to visiting the Down House, which is located just a few miles South East of London, I went to Canterbury, Kent, to attend the International Society of Protistologists (ISoP) annual meeting, at the University of Kent. Coincidentally, back in 1991, as an undergraduate student, I obtained a Diploma in Endangered Species Management from the University of Kent, which offered such certification in partnership with the Jersey Wildlife Preservation Trust (nowadays Durrell Wildlife Conservation Trust)”… Read MORE.
Some History about Darwin Day at the US Congress
Click on image to watch video. Paul Broun (R) “All that stuff I was taught about evolution… all that is lies straight from the pit of Hell”
“…Bill H.Res.41, itself, embodies the never-ending battle against irrationalism, the latter vividly present in the views of those who see evil in truth and menace in the realities discovered by science. Take, for example, last year’s remarks by congressman Paul Broun (R), from Georgia, a physician and member of the House of Representatives Committee on Science, Space and Technology (the very Committee to which the “Darwin Day bill” was referred), who declared: “God’s word is true. I’ve come to understand that. All that stuff I was taught about evolution and embryology and the big bang theory, all that is lies straight from the pit of Hell.”
And Mr. Broun went on, as documented in video watched worldwide: “It’s lies to try to keep me and all the folks who were taught that from understanding that they need a savior.” “You see, there are a lot of scientific data that I’ve found out as a scientist that actually show that this is really a young Earth.” “I don’t believe that the earth’s but about 9,000 years old.” “I believe it was created in six day as we know them.” “That’s what the Bible says.” Read MORE.
Today in EvoLiteracy News: A recent study about the authenticity of the Shroud of Turin caught my attention for two reasons. First, it was published in Scientific Reports, a science-media outlet sponsored by Nature. Second, five authors believed to have tested two “hypotheses” when, in reality, they reconfirmed –indirectly– support for an old thesis (i.e. the Medieval origin of the shroud, 800 years ago) and speculated over a historicity-based idea (i.e. that the shroud originated in Jerusalem, 2000 years ago, and it was later transported to Turin). The study is an exemplar of conceptual error, and it can be used in scientific-methodology courses to educate our students on how to avoid making similar mistakes. – Guillermo Paz-y-Miño-C
Shroud of Turin, Poor Science, and the Persistence of a Myth
I will dedicate some length to examine this study (Uncovering the Sources of DNA Found on the Turin Shroud), which falls apart by itself when attempting to combine modern DNA analysis with a silly, pseudoscience project: to find scientific evidence to justify believing in a myth. The concerning part is that five authors, Barcaccia et al., were fortunate to publish the research (October 5, 2015) in Scientific Reports, a fairly known science-communication venue.
Negative of the Shroud of Turin printed as positive 1898
The team examined “two” hypotheses, one scientific, the other a historicity supposition. The scientific thesis –for which Carbon dating has provided unequivocal evidence since 1988/9— is that the Shroud of Turin is a fake, a fabrication of Jesus’ body’s imprint on ordinary linen traceable to the 13th and 14th centuries, specifically to 1260-1390 (C-dating is quite accurate). The historical supposition, in contrast, suggests that the shroud must have come from the years 30 or 33, of modern times, which makes it a 2000-year-old relict. The latter has no scientific sustain. But the authors build up –in a technical report format– the argument that their molecular analyses bring support to “both” hypotheses (Medieval origin and Jesus’ timeline). This is not the case. In fact, it is impossible to merge both proposals because the shroud is 800 years old, while the Jesus-person tale –if real– is three times older. Therefore, there is an irreconcilable mismatch in time between the two views.
Readers can judge the study for themselves (at this link), but I prefer to summarize the reality of its findings –while avoiding the authors’ convoluted logic– as follows:
First, Barcaccia et al. admit that the Carbon-dating piece of evidence –a crucial element in this case– demonstrates that the Shroud of Turin originates in Medieval times (statistical confidence 95 percent). That is, we are pretty sure that the linen does not come from 2000 years ago. The shroud is one third the age it is supposed to be under the historicity hypothesis. Therefore, everything else upon which the authors speculate in the study is irrelevant. The Barcaccia et al. conjectures, no matter how the investigators spin them, are inconsistent with the Carbon-dating clock: 800 years old.
Second, the shroud is contaminated with plant material (particularly pollen, which can be detected via chloroplast DNA analysis) belonging to plant species from, almost, all over the world (see figure below). Because the shroud’s linen is 800 years old, the contamination must have occurred during recent times.
Plant DNA species found on the Shroud of Turin. Schematic overview of the centers of origin of plant taxa identified in samples of the shroud. Image source Barcaccia et al. 2015.
Third, the linen of the shroud is also heavily contaminated with human DNA (detected via mitochondrial DNA profiling; see figure below), which belongs to multiple individuals, from multiple ethnic backgrounds and, therefore, from multiple geographic locations (Europe, West Asia, Northeastern Africa, Arabia, Middle-East, all the way down to India –not from the Americas). Again, because the shroud is 800 years old, human contact with it must have occurred during recent times.
Human mtDNA haplogroups found on the Shroud of Turin. Schematic overview of the current geographical distribution of human mtDNA haplogroups and sub-haplogroups identified in samples of the shroud. Image source Barcaccia et al. 2015 (click on it to enlarge).
Fourth, and this is not a trivial finding, although the authors report it as extraneous, the shroud is also contaminated with DNA from the bird Southern Grey Shrike (Lanius meridionalis), distributed in Southwest Europe, Northern Africa, the Near East, and Southwest Asia. And also with genetic traces of a marine Nemertine worm (Cerebratulus longiceps), from the Northern Pacific Ocean (species description from Alaskan samples, 1901). Once more, as it becomes crucial to remind the readers, because the shroud is 800 years old, bird- or marine worm-DNA must have contaminated the fabric during the past eight centuries.
Fifth, despite these findings, the authors bend over backwards, put aside the age of the shroud and free fall into speculating that if we –somehow– eliminate the noise of the overwhelming DNA contamination, we can “parallel” the genetic patterns of human DNA contamination with the historical path taken by the shroud’s carriers, allegedly from Jerusalem to Turin. A sort of “spatial” retracing of the shroud’s migration. That is, an attempt to find in the genetic markers –which in reality only inform us about the diverse geographic sources of the DNA contaminants– the evidence in support for the historicity hypothesis: Hhthat the linen has traveled from Jerusalem to Turin, and via human hand-to-hand (with occasional bird or marine worm intervention) along an envisioned route. And this way of reasoning –by Barcaccia et al.– is so troubling, because it is a blunt Type One Error, a conceptual violation of scientific scrutiny. It picks and chooses what type of contaminant DNA is more informative than other (when all of them are, in principle, contaminants of a cloth one-third the age it is supposed to be), in order to fit the authors’ wish to force support for the historicity hypothesis. And this is done in plain sight of an 800-year young cloth or, in fact, a fake 2000-year-old-wanna-be relic.
“…To be authentic, the Shroud of Turin must be 2000 years old. It is not. Carbon dating places it only eight centuries ago, in Europe. And there is no evidence to link the shroud to Jerusalem, and to the years 30 or 33.”
Sixth, what Barcaccia et al. seem to have found, after cleaning some of the noise in the multiple genetic contaminations (i.e. by discarding some contaminants and keeping “informative” DNA sequences) is an artifact, a byproduct effect of the spatial source of the contamination, that gives the impression of a geographic pattern of migration of the shroud (from Sacred Land to Turin). In reality, it is a pattern of the source of the contamination (not a reflection of the origin of the shroud), consistent with the history of human-worshipers’ approaches to the linen, to touch it. This actually explains the apparent absence of human DNA contaminants from the Americas in contrast to the more-likely-contaminants to be found: the people living nearby the shroud, from the years 1200-1300s onwards. And this is a more parsimonious explanation to the Barcaccia et al.‘s speculations. The authors could have just rationalized over a 2×2 table (see figure below) when designing the tests for: Medieval-origin hypothesis (spatial and temporal evidence) and historicity hypothesis (spatial and temporal evidence). In other words: Medieval-origin hypothesis (Europe and 800 years) and historicity hypothesis (Jerusalem and 2000 years). It becomes obvious that the authors cannot combine, in any manner, one hypothesis with the other because the proposals do not match in time (the time dimension). Therefore, any spatial speculation of consistency, as Barcaccia et al. propose, is senseless. More so when the spatial argument for “possible parallelisms” between hypotheses relies on the capricious use of DNA-contaminants and the exclusion of the timeframe disparity (800 vs. 2000 years). Poor science, indeed.
To be authentic, the Shroud of Turin must be 2000 years old. It is not. Carbon dating places it only eight centuries ago, in Europe. And there is no evidence to link the shroud to Jerusalem, and to the years 30 or 33. A: spatial evidence for the Medieval origin hypothesis vs. the Historicity hypothesis; European art techniques used to fake the shroud have been confirmed vs. the genetic contaminants used to speculate that the shroud may come from the Sacred Land. B: Temporal vs. spatial evidence; radio Carbon-dating places the shroud 800 years ago, which is inconsistent with the hypothetical Jerusalem origin. C: Temporal vs. spatial evidence; the hypothetical 2000-year-old age of the shroud is inconsistent with the Medieval European art techniques used to fake it 800 years ago. D: Temporal vs. temporal evidence; the 2000 years of age required for the hypothetical Jerusalem origin of the shroud is inconsistent with the actual 800 years of age determined by Carbon-dating. – GPC
CONCLUSION: Barcaccia et al. study is another, among many, in-text acknowledgments that the shroud of Turin is a Medieval fake, only 800 years young (C-dating) and not a 2000-year-old relic (as demanded it to be in any test of the historicity hypothesis). Reality occurs in a space-time context, yet the authors chose to put time aside to facilitate survival of the historicity tale. The shroud is heavily contaminated with plant, human, bird and marine worm DNA, a phenomenon enhanced exclusively during the past eight centuries, during which the linen has had increasing exposure to people –and their debris, including critters’ goo– from Europe and nearby locations (not to mention the rest of the world). The genetic spatio-temporal mapping presented by Barcaccia et al. reflects a pattern of contamination by humans of multiple ethnicities, consistent with convergent traveling, from diverse locations, in the direction to the object, the shroud, possibly to worship it. If anything, this paper documents the spatial distribution of human contamination of the linen from likely-geographic locations. It fails to provide any evidence in support of the historicity hypothesis (due to the time dimension incongruity), and it actually strengthens the notion that the shroud of Turin is a Medieval fake. The study by Barcaccia et al. is exemplar of how far poor judgment can go, and how belief in supernatural causation disrupts, distorts, delays or stops (the 3Ds + S cognitive effects of illusory thinking) the acceptance of scientific evidence before the very eyes of the investigators, and with their own data. – GPC
Negatives (left and right images) of the Shroud of Turin. The cloth is about 14 x 4 ft.
VIDEO: Click on image below to be redirected to NatGeo video Remaking The Shroud, in which techniques for faking it are discussed, and some modern-technology tests shown, including UV light and 3-D imaging.
Today in EvoLiteracy News: Louder Vocalizations in Howler Monkeys Correlate with Small Testicles. The significance of this study is the simultaneous test of three hypotheses (vocal competition, environmental adaptation and size exaggeration) and the elegant documentation of an “evolutionary trade-off” in which vocalizations (rather than the usual morphological traits) are involved. Enjoy! – GPC
In 1871, Darwin suggested that the vocal organs of male Mycetes (Alouatta) –howler monkeys– have been sexually selected. What for? Darwin implied that, in sexually dimorphic species, females prefer and mate with males possessing conspicuous ornaments (e.g. large antlers, bright coloration, elaborate vocalizations), which convey information about fitness.
Darwin wrote: “…The vocal organs of the American Mycetes caraya [Alouatta caraya] are one-third larger in the male than in the female, and are wonderfully powerful. These monkeys in warm weather make the forests resound at morning and evening with their overwhelming voices. The males being the dreadful concert, and often continue it during many hours, the females sometimes joining in with their less powerful voices… Whether most of the foregoing monkeys have acquired their powerful voices in order to beat their rivals and charm the females, or whether the vocal organs have been strengthened and enlarged through the inherited effects of long-continued use without any particular good being thus gained, I will not pretend to say; but the former view, at least in the case of [another primate] the Hylobates agilis [the gibbon*], seems the most probable…” (The Descent of Man and Selection In Relation to Sex, 1871).
[*Darwin highlights that the gibbon (H. agilis) “…is remarkable from having the power of giving a complete and correct octave of musical notes, which we may reasonably suspect serves as a sexual charm…]
A new study published in Current Biology explains that hyoid bones (crucial for roars in Alouatta) are highly sexually dimorphic, vary among species and with greater sexual dimorphism in species with larger hyoids. The study examines a phenomenon called “evolutionary trade-off;” a situation in which the enhanced expression of one trait (hyoid bone) is directly associated with the diminished expression of another (size of testicles). The assumption is that both traits are costly for an organism to express and, therefore, the trade-off resides in allocating resources to only one of them, for optimization.
Here is the story: All species of howler monkeys have a modified larynx with an enlarged hyoid bone, which works as a resonance chamber. Howls are energetically expensive to produce (link to video below). Now, what is the connection between the monkeys’ vocalizations and the size of their testicles? The study, authored by Dunn et al. (nine co-authors), explores three hypotheses: vocal competition (i.e. acoustic signals announce presence during defense of territories), environmental adaptation (i.e. hyoid bones are important for sound frequency adjustments in dense forests), and size exaggeration (i.e. voluminous hyoids work as enhancers of the acoustic impression of body size conveyed by roars).
RIGHT: Skeleton of a male Alouatta in which the hyoid bone is clearly visible –throat region. LEFT: two images of the hyoid bone showing its chamber-like structure (top) and smooth surface and elongated shape (bottom). The hyoid is very robust (made mostly of compact bone tissue).
Dunn et al. found support for the vocal competition and size exaggeration hypotheses, but not for the environmental adaptation. Here are some of the authors’ generalizations:
(1) males of various species, who live in larger groups of males, have smaller hyoid bones (panel A in figure below) and larger testicles (panel B), the latter an adaptation for sperm-sperm competition (the trade-off: because both roars and sperm production are expensive, only one trait is conspicuously expressed);
(2) males with large hyoid bones have smaller testicles (panel C in figure above; the trade-off again; these monkeys usually live in single-male-and-many-females groups);
(3) males with larger hyoid bones vocalize at low(er) frequencies (panel D in figure above; size exaggeration hypothesis); and
(4) all howler monkey species produce very low-frequency vocalizations in respect to their body mass and in contrast to other mammals or corresponding body mass (see figure below; also consistent with the size exaggeration hypothesis). – GPC
VIDEO: Take a look at the energetic movement of the thorax while this howler monkey vocalizes; the entire anatomy participates in the roaring. In the rainforest, howler monkeys’ vocalizations can heard from kilometers away.








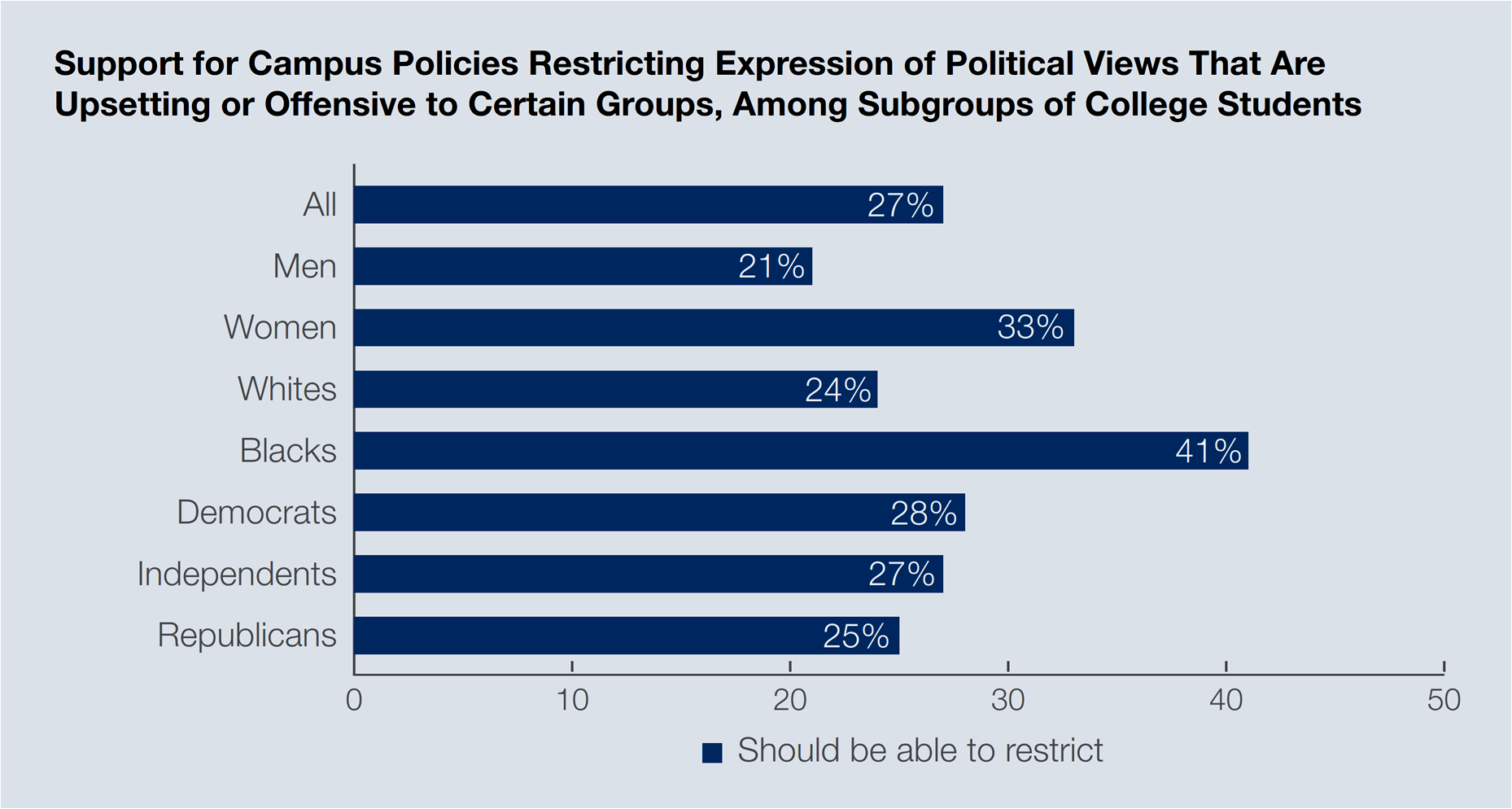
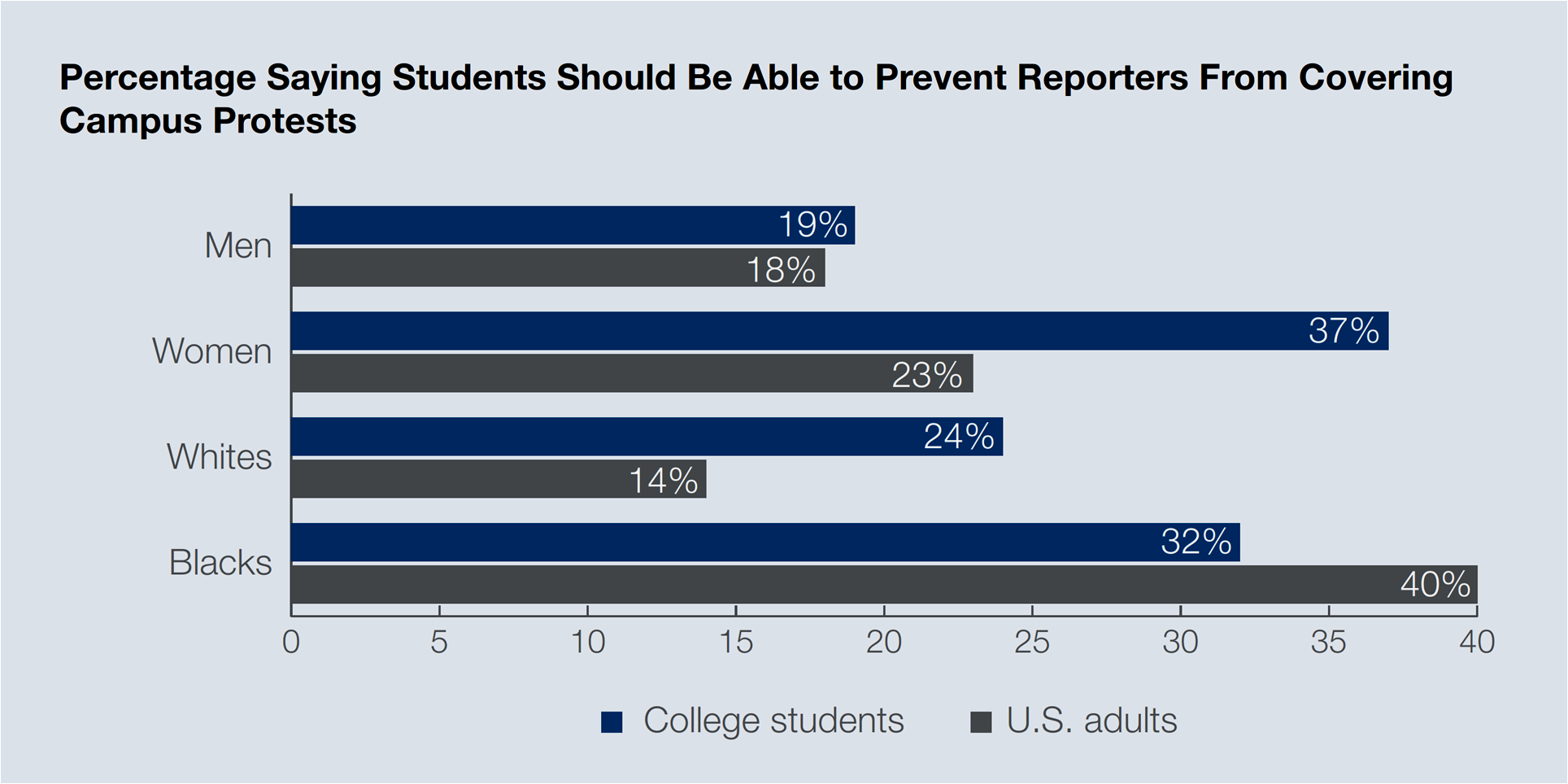




















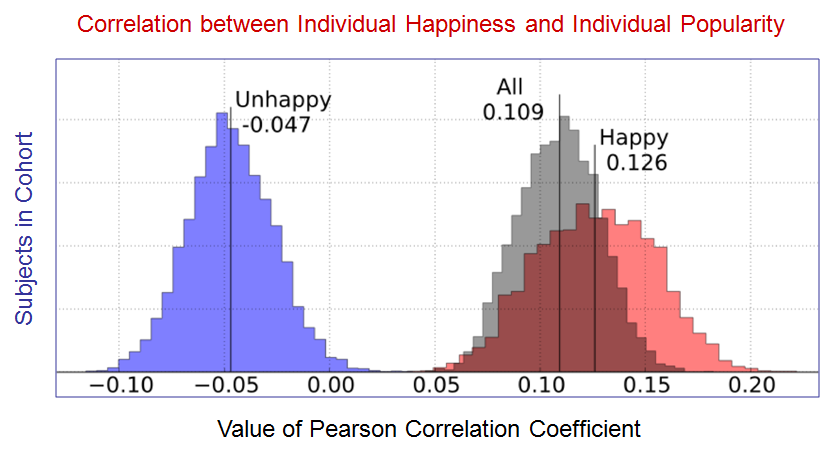




















You must be logged in to post a comment.