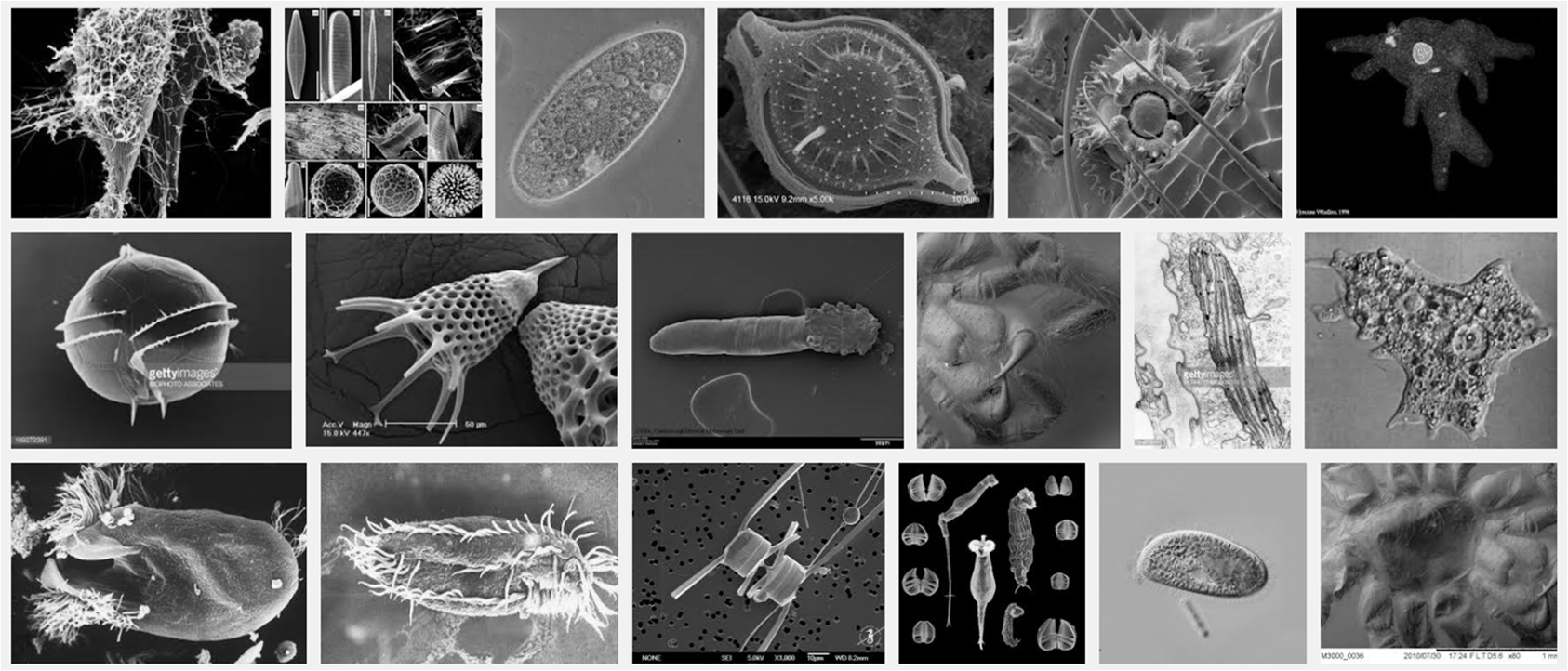
Our latest review on clone-clone discrimination/recognition in Entamoeba species has been featured on the cover of the Journal of Eukaryotic Microbiology (JEUK-MIC, Volume 66, Issue 2, March-April 2019). This is the second time our work has made it to the cover of JEUK-MIC (the first time was back in 2012, coincidentally in the March-April Volume 59, Issue 2). Below are the links to the 2019 article, as well as the abstract and caption to the cover image.
Entamoeba histolytica is one of the least understood protists in terms of taxa-, clone- and kin-discrimination/recognition ability. Yet, the capacity to tell apart same or self (clone/kin) from different or non-self (non-clone/non-kin) has long been demonstrated in pathogenic eukaryotes like Trypanosoma and Plasmodium, free-living social amebas (Dictyostelium,Polysphondylium), budding yeast (Saccharomyces), and in numerous bacteria and archaea (prokaryotes). Kin discrimination/recognition is explained under inclusive fitness theory; that is, the reproductive advantage that genetically closely related organisms (kin) can gain by cooperating preferably with one another (rather than with distantly related or unrelated individuals), minimizing antagonism and competition with kin, and excluding genetic strangers (or cheaters = non-cooperators that benefit from others’ investments in altruistic cooperation). In this review, we rely on the outcomes of in vitro pairwise discrimination/recognition encounters between seven Entamoeba lineages to discuss the biological significance of taxa-, clone- and kin-discrimination/recognition in a range of generalist and specialist species (close or distantly related phylogenetically). We then focus our discussion on the importance of these laboratory observations for E. histolytica‘s life cycle, host infestation, and implications of these features of the amebas’ natural history for human health (including mitigation of amebiasis).
“…Belief is powerful, it disrupts, distorts, delays or stops the comprehension and acceptance of scientific evidence. Scientists call this phenomenon the ‘3Ds+S’ cognitive effects of illusory thinking. Now, the best tonic against its infectious sequels was discovered long ago; it consisted, still does, on proper healthcare education for all. And, in contrast to misleading and unwarranted ‘paramedicine,’ the side effects of widespread science education will always be cheaper, plus save, for sure, some lives…”
By Guillermo Paz-y-Miño-C
Cancer hides or thrives in our bodies. Someone we know, close or distant, is destined to die because of it. And although therapies continue to improve thanks to scientific advances, diverse cancers persist and it might take decades, if ever, to fully manage them.
In the United States, breast, prostate, lung and colorectal cancer are the most prevalent, and chemotherapy, radiotherapy, surgery, and/or hormone therapy the usual treatments. Yet, more than half of the patients with cancer opt for “complementary medicine” to improve, as they believe, their quality of life and survival.
But, do herbs and botanicals, vitamin and mineral supplements, probiotics, traditional medicines, homeopathy, naturopathy, acupuncture, chiropractic or osteopathic manipulation, massage, prayer, reflexology, energy medicine, or special diets have an actual impact on prolonging cancer patients’ lives? The short answer seems to be no.
Physicians from the Yale School of Medicine have just published the study “Complementary Medicine, Refusal of Conventional Cancer Therapy, and Survival Among Patients with Curable Cancers” in the Journal of the American Medical Association (JAMA). From a data set of 1.9 million individuals, gathered between 2004 and 2013 and stored in the National Cancer Database, the researchers extracted a representative sample of cancer patients whom opted for complementary medicine (CM) versus those exposed to conventional cancer treatment (CCT).
The study was straightforward. It aimed at identifying and comparing survival rates between CM and CCT groups.
Patients exposed to complementary medicine had a greater risk of death than those under conventional cancer treatment. In fact, only 82 percent of the CM patients versus 87 percent of the CCT patients survived during a 5-year monitoring lapse since they were first diagnosed with the condition. The trend was noticeable in women suffering breast cancer, with only 85 percent of the CM patients versus 90 percent of the conventionally-treated patients surviving since diagnosis (also during a 5-year follow up).
According to Skyler Johnson, Henry Park and Cary Gross, authors of the study and fellows at the Department of Therapeutic Radiology, as well as the Cancer Outcomes, Public Policy, and Effectiveness Research Center at Yale, the general risk of death associated with complementary medicine was primarily linked to the patients’ refusal to receive: surgery (7 versus 0.1 percent refusal between the CM versus CCT groups, respectively), chemotherapy (34 versus 3 percent), radiotherapy (53 versus 2 percent), and/or hormone therapy (34 versus 3 percent).
Quite interestingly, patients in the complementary medicine group were more likely to be young, women, have breast or colorectal cancer, belong to high socio-economic cohorts, have private medical insurance, high-school education, and reside in the Intermountain West or Pacific West of the United States (where alternative-medicine schools are common, protected by state legislation).
In essence, the Yale study concluded that if patients went for unconventional cures to fight cancer, rather than scientific medicine, they had higher risk to die and do it earlier. Complementary medicine did not help.
As David Gorski, member of the Department of Surgery at Wayne State University School of Medicine and the Department of Oncology at the Barbara Ann Karmanos Cancer Institute (both in Michigan), already alerted back in 2014 when compiling a comprehensive review for the journal Nature “the vast majority of ‘integrative’ [complementary oncology] treatments [were] supported by little, if any, scientific evidence.” He pointed out, with concern and irony, “therein lies a key problem with integrative oncology. The less ‘alternative’ the intervention, the more it resembles conventional oncology; the more ‘alternative’ the intervention, the more it resembles the quackery from which integrative oncologists rightly distance themselves.”
Why do patients opt for unscientific methods to battle cancer? There are multiple reasons, and only one of them has to do with “hope,” trust on a possibility (the “alternative cure”) beyond the “conventional scientific wisdom,” one that might work and, if not, at least, it won’t hurt. There is always a friend or a relative that recommend “holistic cures” to somebody they love. But the Yale study demonstrates that such paths can indeed be harmful: remember that they were associated with higher risk of dying and doing it earlier among patients choosing “complementary practices.”
“…Academics ‘are trapped; compelled to participate in activities they find distasteful,’ including the intricate world of scientific publications, which involves a range of journal publishers, editors, book producers, open-access periodicals, for-profit series, online-journals and other venues to disseminate research; plus, of course, individuals —charming, powerful or both— that free ride at the expense of others’ work…”
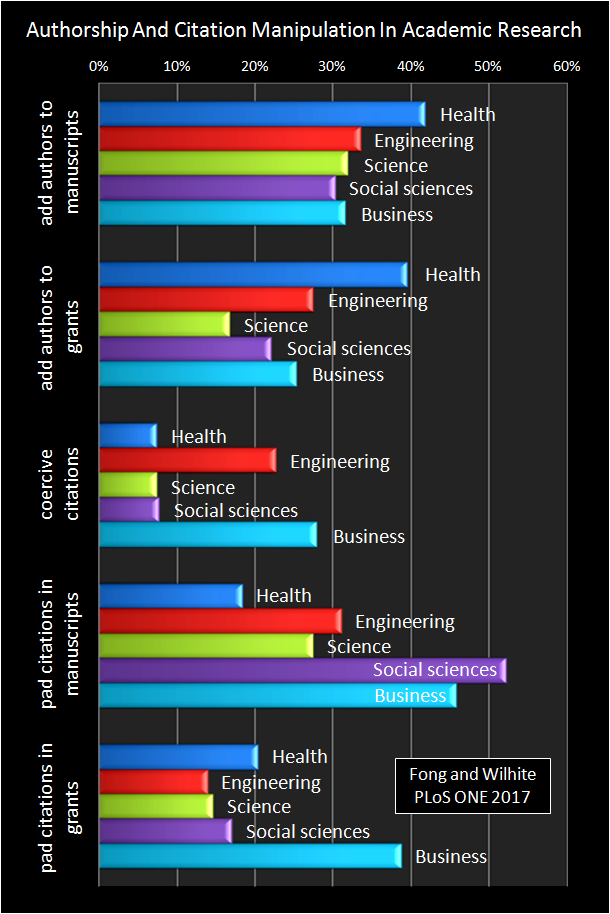
In an article just published in PLoS ONE, December 2017, Eric A. Fong and Allen W. Wilhite, researchers at the University of Alabama in Huntsville, document three practices in academia: (1) the inclusion of “honorary authors” in scientific publications (i.e. the addition of individuals to manuscripts as authors, even though they have contributed little, if anything, to the actual research), (2) coercive citations (i.e. when editors direct authors to add citations to articles from the editors’ journals ‒arguably to boost the journal’s citation index), and (3) padding (i.e. when authors add superfluous citations to a paper in an attempt to increase its chance for publication). Fong and Wilhite surveyed 12,000 scholars from 18 disciplines (i.e. health-care, engineering, science, social sciences and business) at diverse universities in the United States.
The specific fields of specialization of the interviewed researchers were: medicine, nursing, biology, chemistry, computer science, mathematics, physics, engineering, ecology, accounting, economics, finance, marketing, management, information systems, sociology, psychology, and political science. The Fong and Wilhite’s study is comprehensive and I suggest the reader to look at the original publication for details. In the figure below, I simply share the statistical trends that summarize the research. I have adapted an image from the PLoS ONE article (open access journal) to make it more appealing to a non-specialized audience.
In the image, note the following: (1) Adding honorary authors to manuscripts was common practice among 1 in every 3 of the surveyed scholars; it reached +40% in the health-care professions. (2) About 1 in every 5 scholars added such authors to a grant proposal; and, again, about 40% was typical in the health-care professions. (3) Coercive citations were common in +5% of the scholars in science, and beyond 20-25% in engineering and business. (4) The practice of padding a scientific article with irrelevant citations was common among, at least, 1 in every 5 authors, and particularly evident among those in science and engineering (around 30%), and +45-50% in business and the social sciences. Finally, (5) padding in grant proposals was common in +10-20% of all disciplines, and up to ≈40% in business.
The responders to the survey acknowledged that the main reason for adding honorary authors in their manuscripts was the relationship of director/authority of the honorary author in respect to the “real authors” of the paper (+20% of the responders thought that way); others included as co-authors their mentors, colleagues, individuals-for-reciprocity-reasons, for their reputation, or for funding. However, 60% of all responders added honorary authors to their grant proposals simply due to the latter’s “reputation,” and, thus, to increase the chances of getting the projects funded.
Interestingly, between ≈60% and +90% of all responders disapproved of the coercive citation practice across disciplines. But keep in mind that, despite the quantitative approach to the study, the responses were also based on perception, i.e. the researchers’ insight about honorary authors, coercive citations or padding in scientific publications and grant proposals.
Fong and Wilhite concluded that:
“…there is a significant level of deception in authorship and citation in academic research and while it would be naïve to suppose that academics are above such scheming to enhance their position, the results suggest otherwise. The overwhelming consensus is that such behavior is inappropriate, but its practice is common. It seems that academics are trapped; compelled to participate in activities they find distasteful…”
“…misattribution, spans the academic universe. While there are different levels of abuse across disciplines, we found evidence of honorary authorship, coercive citation, and padded citation in every discipline we sampled. We also suggest that a useful construct to approach misattribution is to assume individual scholars make deliberate decisions to cheat after weighing the costs and benefits of that action. We cannot claim that our construct is universally true because other explanations may be possible, nor do we claim it explains all misattribution behavior because other factors can play a role. However, the systematic pattern of superfluous authors, coerced citations, and padded references documented here is consistent with scholars who [are] making deliberate decisions to cheat after evaluating the costs and benefits of their behavior…”
To close: In my (our) own experience, I (we) have never included honorary authors in my (our) publications, or have never been an honorary author of a paper. But some colleagues have asked me (us), more than once, why have I (we) included such and such person in that or that paper, assuming that that individual did not deserved it. My (our) rationale has always been that if a substantial aspect of a manuscript had not been possible to be completed, unless that person had contributed directly or indirectly to the research, that individual ought to be acknowledged as co-author. This includes the very conceptual transformation of a manuscript due to crucial feedback, interpretation of results, and substantial modification of the scope with which an article was put together. — In cases like that, I have expected my name to be included as co-author, but that has happened sporadically (more during the times I was a postdoc and contributed with conceptual, methodological, analytical and copy-editing feedback to graduate students’ dissertations). In various occasions, I have requested to not be included in manuscripts as a co-author; this practice is not unusual among researchers.
In terms of coercive citations, I have never been asked by a journal editor to cite an irrelevant paper with the purpose of contributing to boosting the journal’s citation index (which, by the way, would require hundreds of authors to be simultaneously coerced to cite multiple articles to have a statistical influence on the journal’s performance), although I have been suggested by journal editors to take a look at some studies (published in other journals), of which I was not aware, and that I actually found very helpful to come across, and decided to discuss and cite them in a paper. However, and this is a big “however,” peer-reviewers have attempted to coerce us (Avelina Espinosa and I), more than once, to cite their papers (or their close collaborators’), or papers of their liking, in our studies. In many cases, such papers were irrelevant, or we disagreed fundamentally with them to even give them a citation in our manuscripts. On one occasion, a well known individual in a field insisted that we should cite non-scientific books in our work, and did so with assertive authority (plus specifically stated where in our text we had to acknowledge the merits of the organization with which the individual was affiliated ‒sounds surreal, right?), an issue we later resolved with the journal editor, who agreed with us and considered the suggestion to be imprudent. So, yes, coercive predators do exist and attempt to exert power at will… if you let them. But journal editors are very experienced, for the most part, and tend to not allow such approaches to peer-reviewing.
On grants, we have never included potential honorary recipients to increase our chances of getting funded. But, when attending a national-funders meeting in Washington DC, a few years ago, we were advised to add a specific anchor-individual to our team, otherwise “we will continue to be seen as outsiders” (verbatim) in that specific community of peer-reviewers. Of course, we declined to include that person in our proposals (three of which were not funded by the agency), although we did complete the research and publish the papers (N = 14) that we projected in the proposals to be the outcomes of the projects. Not only that, we were fortunate to publish an academic best-seller-2017-book summarizing all the research and with no strings attached to any honorary contributor.
“…Colleges and universities must lead freedom of speech, nurture the battlefield of ideas, pursue world-competitive standards, deter grade inflation (a historical, pervasive companion of conflict resolution), stand for science and reason, teach the realities of the cosmos and evolution without spiritual justifications, and be the safe spaces where the trigger warning ‘no ignorance allowed’ is respected…” – GPC
[click on subtitle to be redirected to The Standard Times]
“There’s been a trend around the country of trying to get colleges to disinvite speakers with a different point of view, or disrupt a politician’s rally. Don’t do that, no matter how ridiculous or offensive you might find the things that come out of their mouths. Let them talk. If you don’t, you just make them a victim, and then they can avoid accountability.”
“That doesn’t mean you shouldn’t challenge them. Have the confidence to challenge them, the confidence in the rightness of your position. But listen. Engage. If the other side has a point, learn from them. If they’re wrong, rebut them. Teach them. Beat them on the battlefield of ideas.”
These were President Obama’s remarks at the Howard University commencement ceremony, back in May 2016. He highlighted a nowadays generalized concern in academia: that millennials (the generation born between the 1980s and early 2000s) have grown intolerant of any view contrary to their deepest, yet maturing convictions, a phenomenon documented by Gallup in its latest report “Free Expression On Campus.”
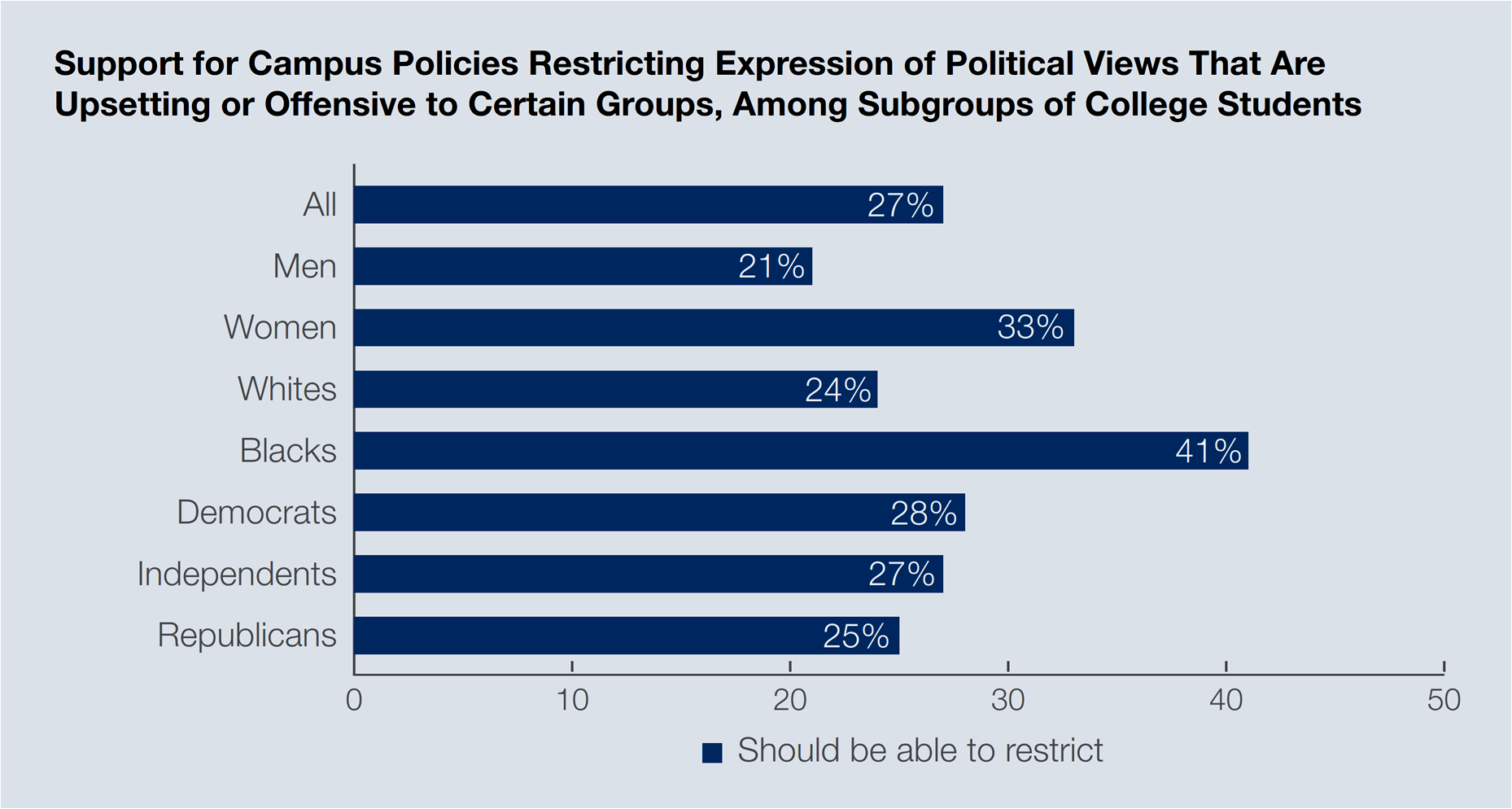
Although “college students believe First Amendment rights remain strong” in the United States, they also hold conflicting views about “shuttering free speech and impeding a free press under certain circumstances.” For instance, even though 81 percent of college students think that freedom of the press is very secure or secure in the country, and 73 percent think likewise about freedom of speech, one in every five college students (22 percent) believe that “to create a positive learning environment for all, it is more important for colleges to prohibit certain speech or expression of viewpoints that are offensive or biased, than to create an open learning environment where students are exposed to all types of speech and viewpoints —even offensive or biased.”
“…boycott actions against free speech may be rooted in capricious interpretations of First Amendment rights by students active and verbal at their institutions…”
But, what is offensive or biased rhetoric? Slurs and language that intentionally seek to hurt or offend (69 percent of students think so), or the wearing of costumes that stereotype racial or ethnic groups (63 percent believe that), or political views that may upset or offend (27 percent of students think colleges should limit such speech). The Gallup data, therefore, implies that boycott actions against free speech may be rooted in capricious interpretations of First Amendment rights by students active and verbal at their institutions.
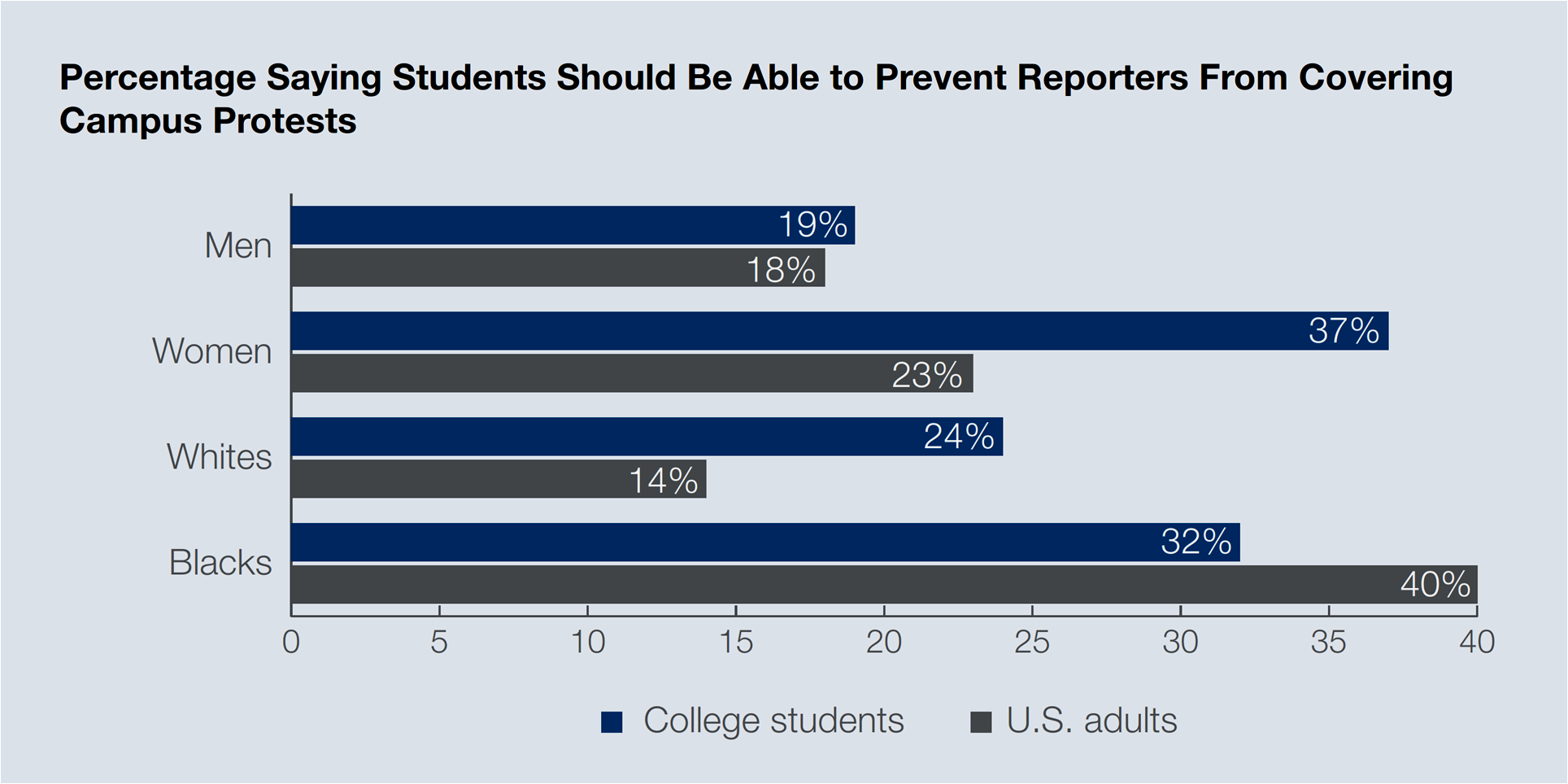
Should the media cover protests held at colleges and universities? One in every four students (28 percent, mean of men + women) opposes reporters’ coverage of demonstrations in campus. This view is noticeable among women (37 percent) and African Americans (32 percent). Gallup calls this overall occurrence “support of free press rights in the abstract” (or “in principle”) but not in practice. Protesters believe the press will be unfair in its reporting (49 percent say this), they assert to have the right to be left alone (48 percent), and want to tell their own version of the story on the internet and social media (44 percent). In fact, 86 percent of the polled students prefer the social media for allowing them more control over the story.
Why has intolerance of free speech become ubiquitous? Only tentative answers exist to this question. Greg Lukianoff and Jonathan Haidt, authors of “The Coddling of the American Mind,” have taken a psychological path to explain how “in the name of emotional well-being, college students are increasingly demanding protection from words and ideas they don’t like.” The ultimate aim, Lukianoff and Haidt state, “is to turn campuses into ‘safe spaces’ where young adults are shielded from words and ideas that make some uncomfortable. This movement seeks to punish anyone who interferes with that aim, even accidentally.”
“…Emotional reasoning, distorted thinking, magnification of events, and catastrophizing incidents seem to be central to the individual and collective minds of those who perceive offense in daily experiences…”
The “impulse” is called “vindictive protectiveness” and, as described by Lukianoff and Haidt, “it is creating a culture in which everyone must think twice before speaking up, lest they face charges of insensitivity, aggression, or worse.” Emotional reasoning, distorted thinking, magnification of events, and catastrophizing incidents seem to be central to the individual and collective minds of those who perceive offense in daily experiences. Thus, via negative filtering, groups develop a culture which focuses almost exclusively on the unconstructive, and this allows for simpleminded demonization.
“…The challenge is to identify objectively what is intellectually or psychologically damaging and what is mild…”
Universities must be the safe spaces where the trigger warning “no ignorance allowed” is respected – GETTY IMAGES
“The great contribution of ‘Measuring the Evolution Controversy’ is the rich content of data and analysis that asks detailed questions about the social, economic and political backgrounds of those who tend to reject evolution vs. those who accept evolution as science. Paz-y-Miño-C and Espinosa deftly analyze their data drawn from institutions of higher learning in the United States and particularly New England —which stands as a microcosm of the rest of the country, and indeed elsewhere in the world. It is their scientific approach to these issues which makes this book stand out as a uniquely original contribution.” —Niles Eldredge, PhD, Curator Emeritus of Paleontology at The American Museum of Natural History, New York.
“Pro-science activists and educators constantly bemoan the resistance to the teaching of evolution in the United States. All of us have anecdotes about encounters with the public, parents and students who are misinformed by their churches, Religious-Right groups, and creationist organizations. Paz-y-Miño-C and Espinosa present hard data that support the anecdotal evidence. They also show that although anti-evolutionism typically begins with religion, it is a multi-faceted problem that intersects with political and cultural ideologies. Gathered through careful research over a period of years, their data will enable scientists and defenders of science education to comprehend the roots of the evolution controversy and counteract resistance to evolution more strategically and effectively.” — Barbara Forrest, PhD, co-author with Paul R. Gross of Creationism’s Trojan Horse: The Wedge of Intelligent Design (2007), and expert witness for plaintiffs, Kitzmiller et al. v. Dover Area School District (2005).
“The sweet spot of this collection of essays is the interface of science, history and literacy. Paz-y-Miño-C is, in essence, a champion of rationalism and a passionate defender of literacy standards. His essays deftly weave hard survey data and memorable turns of phrase with evocative imagery… While the essays in this collection are vast in coverage —from climate change to energy policy, stem cell research, vaccinations and, especially, evolution— a clear underlying theme emerges: [the author’s] goal is no less than to counter, through the lens of history and the majesty of rationalism, social forces that sanction ignorance, celebrate denial and… continue to diminish our global status in the fields of science and technology.” Jeff Podos, PhD, Professor of Biology, University of Massachusetts Amherst, USA.
“Paz-y-Miño-C is a firm believer in evolutionary processes. He would like to see decisions made on the basis of facts, not unsupported opinion. He abhors and fears irrational thinking, especially ‘the views of those who see evil in truth and menace in the realities discovered by science.’ He marvels at the intricacy and diversity of life, and how it came about through natural selection… and is clearly frustrated by the unwillingness of so many to see the beauty and majesty in this view of the world and all that it explains.” – Jan A. Pechenik, PhD, Professor of Biology, Tufts University, USA, author of The Readable Darwin: The Origin of Species, as Edited for Modern Readers.
This morning, I learned from Avelina Espinosa that our symposium articles on Kin-Discrimination in Protists have been featured on the latest issue of the Journal of Eukaryotic Microbiology (May-June 2016). I posted a report about this topic a few weeks ago, when the papers became available online as “early view.” Here are page updates about these publications and a view of the journal cover (i.e. several developmental stages of the social amoeba Dictyostelium discoideum; photo by Usman Bashir):
To remind the readers, these articles summarize outcomes of the symposium “Evidence of Taxa-, Clone-, and Kin-discrimination in Protists: Ecological and Evolutionary Implications,” which took place at the VII European Congress of Protistology, organized last summer in partnership with the International Society of Protistologists (ECOP-ISOP 2015), and hosted by the University of Seville, Spain.
The complete report about the three articles is available at Kin Recognition or Kin Discrimination in Single-Celled Organisms – Protists, which appeared on April 4, 2016, and was featured at EvoLiteracy. As suggested in that report, readers might also benefit from exploring the initial paper that inspired the organization of the ECOP-ISOP 2015 symposium:
Can single-celled organisms distinguish between same (self or clone) and different (diverse clones)? Can they tell apart kin from non-kin? Can they “learn” to discriminate or recognize members of their own versus others’ cellular lineages? Answers to these questions are provided in three articles (first, second, and third) just published (early view) in the Journal of Eukaryotic Microbiology. The studies summarize outcomes of the symposium “Evidence of Taxa-, Clone-, and Kin-discrimination in Protists: Ecological and Evolutionary Implications,” which took place at the VII European Congress of Protistology, organized last summer in partnership with the International Society of Protistologists (ECOP-ISOP 2015), and hosted by the University of Seville, Spain. I participated at this Symposium and co-authored two of the trilogy papers. But my purpose here is to bring attention to the topic “kin discrimination” or “kin recognition” in unicellular organisms, an area of research that has made much progress in the past fifteen years. There are some technicalities in the text below, but I ask the readers to make an effort and try to understand the broad themes, while being patient with the details. Enjoy! – Guillermo Paz-y-Miño-C.
The unicellular eukaryote Entamoeba invadens (a protist) colored with Green or Red markers. When the Green and Red cells fully aggregate in mixed assemblages, they look yellow under the microscope, an indication that the amebas are able to distinguish members of the same clone, without being affected by the coloration. When grown in the laboratory with non-clone members, the amebas only aggregate with same-clone cells, an indication of preference to group with themselves. Photo courtesy of A. Espinosa 2016.
Readers unfamiliar with “protistan” biology might benefit from grasping some terminology (i.e. protist, prokaryote, eukaryote, clone, multicellularity, kin recognition/discrimination), which I explain next by answering simple questions. For those trained in biology, it might be fine to skip the first five subtitles, below, and move on directly to “From multicellular to unicellular: a round trip:”
What is a protist?
Remember that some organisms are made of single cells. Yes, the entire organism is a cell. For example, bacteria (like the E. coli that lives in the human gut) or amebas, which are also microscopic and can be found in a pond. However, one of the main differences between a bacteria and an ameba is that the former has no nucleus, in contrast to the latter that has it. For that reason, bacteria are called prokaryotes (pro = “before” or “prior to” or, in this case, no nucleus) and amebas are called eukaryotes (eu = “well” or “good” or, in this case, with nucleus). However, many other organisms have nucleated cells (and are, therefore, eukaryotes), like a frog, an orchid, a chimpanzee, a maple tree, or a Blue Whale. The term “protist” applies to unicellular eukaryotes, while the amphibian, the plant, or the aquatic mammal are called multicellular eukaryotes, since their bodies are made of billions of nucleated cells.
Now, the term “protist” is tricky because it includes extremely diverse organisms (which earliest ancestors likely emerged two billion years ago), and over the years scientists have realized that many exceptions exist of organisms that, although are not always unicellular in their life cycles, they are (or should be considered) protists. One of them is the social ameba (or amoeba), Dictyostelium, which forms multicellular assemblages (moving “slugs;” watch video) during its life stages, and also lives in nature as a single-celled, free ranging ameba. To learn more about the ambiguities intrinsic to the term “protist,” and for more examples, go here.
What is a clone?
Because unicellular organisms reproduce, in many cases, via simple cell divisions, the resulting progeny is often identical to the parental cell. A single ameba, for example, can give origin to 2 identical daughter cells, which, in turn, can generate 4 cells (i.e. 2, 4, 8… see cartoon on the right). The resulting thousands, or millions, of cells are “clones” (identical copies of each other). Now, during so many cell divisions, mutations that change the sequence of DNA can –and do– occur, making the descendant cells gradually different from the parental ones. Over time, maybe hundreds of years, a cell line could change enough to become a different clone. Moreover, after thousands or millions of years, different species of amebas can emerge, each distinctive from one another and from the species that gave them origin in the past, a phenomenon known as speciation.
What is multicellularity?
Volvox (a green-algae protist) forms large sphere-colonies made of hundreds, or thousands, of individual cells (see daughter colonies inside). Click on image for source.
Perhaps a more didactic question here is ‘where do multicellular organisms (i.e. frogs, orchids, chimps, maple trees, blue whales) come from’? A first, quick answer is that all multicellular organisms known today originated –at some point– in ancient assemblages of unicellular ancestors (traceable to billions of years ago). However, a more cautious answer is that we have a fragmentary understanding about how primitive single-celled eukaryotes took the path (here I mean driven by natural selection) toward permanent associations in immense cellular cooperatives, which we now call multicellular organisms (note that scientists consider the advent of multicellularity a “major evolutionary transition” in the history of Earth). And that is why studying modern protists, like gregarious amebas (in the genus Entamoeba), or facultative social amebas (i.e. not always social, but in response to environmental circumstances), like Dictyostelium, can give us hints about how multicellularity originated. This particular topic is discussed in the three articles published in JEUKMIC (first, second, and third), and to which I refer in this post. But before I get into that, take a look at the image of Volvox (inset). The organism Volvox is a green-algae protist, which forms large sphere-colonies made of hundreds, or thousands, of individual cells. Inside these spheres, daughter colonies develop and, when they mature, the parental spheres bursts and the descendant colonies are released into the aquatic environment, where they continue to grow and proliferate. Today’s multicellular aggregations of protists, like Entamoeba, Dictyostelium or Volvox, give us clues about how multicellularity might have originated in ancient Earth. Moreover, they are good model-systems to study kin recognition or kin discrimination (which include an organism’s skills for grouping and cooperating with the right partners, and behaving altruistically toward them) in the context of the origin and evolution of multicellularity.
What is kin recognition or kin discrimination?
“The ‘field of kin recognition’… has no consensus on definitions or proposed mechanisms, possibly due to the vast diversity of life histories across organisms and their phylogenetic complexities…”
NASA’s twin astronauts Scott, left, and Mark Kelly. Photo: Tony Cenicola. In humans, identical twins are the only natural “clones;” their genetic relatedness is equal to 100% (represented by r = 1.0). However, the rest of us are related to our siblings only by 50%, or r = 0.5. Our relatedness with our parents is the same, r = 0.5. Can the reader tell why? If so, here is a question: what would be your genetic relatedness with an uncle/aunt, or with a grand parent, or with a second cousin?
This topic can be a bit confusing. However, in one of the articles (the first one listed below), the authors explain why: “…The ‘field of kin recognition’… has no consensus on definitions or proposed mechanisms, possibly due to the vast diversity of life histories across organisms and their phylogenetic complexities (here, phylogeny means ancestry, somewhat analogous to genealogy, not of your own family, but rather of distinctive species or kinds of organisms grouped in distinctive categories). [The authors] refer to “recognition” as an organism’s ability to identify kin [family members] versus non-kin [members of another family]; in addition, [the authors] use the term “discrimination” as the capacity to distinguish one clone from another. Because [the authors] discuss instances of taxa- [taxa = in this particular case means species], clone-, and kin-discrimination/recognition in single-celled organisms capable of both discriminating between same and different, and discriminating/recognizing among clones of distinctive [degree of genetic relatedness, like, for example, values of r less than 1.0], [the authors] use these terms together…”
Why is it relevant to study kin recognition or kin discrimination in single-celled organisms?
To answer this question, I will borrow, again, text from the first article: “…Multicellularity is a major evolutionary transition in which single-celled organisms switched from living individually to permanent assemblages. It is possible that multicellularity originated —more than once— in clonality, via a gradual aggregation of closely related cells, capable of recognizing one another by means of chemical cues, and which lived consistently in intimate proximity and benefited from specialized division of labor (i.e. distinctive tissues and organs with given functions). Such specialization included the full allocation of soma-reproduction [soma = the entire body of an organism] to a small population of cells within the soma, the gametes [i.e. ovules, sperm]… Protists are central to the reevaluation of the theoretical framework and concepts in the field of kin recognition, and to research about the origins and evolution of multicellularity...”
From multicellular to unicellular: a roundtrip
The first article (by Paz-y-Miño-C and Espinosa) is a concise review on “Kin Discrimination in Protists: From Many Cells to Single Cells and Backwards.” In it, the authors summarize the current understanding of the genetics of kin discrimination/recognition in unicellular Eukaryotes, and they do it historically by going back in time, to Darwin and his Origin of Species (1859; Darwin speculated* that selection may be applied to the family; kin discrimination/recognition rely on kin-selection theory), and the influential 1960s, when the modern field of kin recognition was, arguably, born. Here is a simplified version of the first article’s abstract:
“During four decades (1960s to 1990s), the conceptualization and experimental design of studies in kin recognition relied on work with multicellular eukaryotes, particularly invertebrates and vertebrates, and some plants. This pioneering research had an animal behavior approach. During the 2000s, work on taxa-, clone- and kin-discrimination and recognition in protists produced genetic and molecular evidence that unicellular organisms could distinguish between same (self or clone) and different (diverse clones), as well as among conspecifics of close or distant genetic relatedness (Table 1, below). Here we discuss some of the research on the genetics of kin discrimination/recognition and highlight the scientific progress made by switching emphasis from investigating multicellular to unicellular systems (and backwards). We document how studies with protists are helping us to understand the microscopic, cellular origins and evolution of the mechanisms of kin discrimination/recognition and their significance for the advent of multicellularity...”
[Click on Table 1, below, to enlarge]
Readers might find the following excerpts from this article quite intriguing, e.g. “learning” in unicellular organisms (is that possible?):
“…In 1899, H. S. Jennings wrote: ‘Paramecium… an animal that learns nothing, that exercises no choice in any respect, that is attracted by nothing and repelled by nothing, that reacts entirely without reference to the position of external objects, that has but one reaction [movement –watch video below] for the most varied stimuli… can hardly be said to have made the first step in the evolution of mind, and we are not compelled to assume consciousness or intelligence in any form to explain its activities.”
Above: movement behavior in Paramecium. This video is 13-min long and shows various types of Paramecium. If you watch it for a couple of minutes, it shall give you an idea about how these ciliateslook like and move.
“Except for mind, consciousness and intelligence, which are not prerequisites for kin discrimination or recognition (both can also operate in a reflex manner: stimulus-response), Jennings was mistaken about his entire characterization of Paramecium. Since the early 1900s, sensitization, trial-and-error learning, and classical or operant conditioning (relevant attributes among some of the multicellular eukaryotes that learn to recognize kin) have been documented in Paramecium; [including] micro-tube-escape swimming behavior via discrimination learning (1910s), habituation to approach baited and un-baited targets using bacteria as food-reinforcer (1950s), and swim-approach behavior toward mild-electrically-charged fields in learning discrimination tasks using positive and punishment reinforcements (2000s).”
And the authors add: “…But, to our knowledge, there is no direct, experimental evidence that protists can rely specifically on sensitization (i.e. the enhancement of a response to an incremental exposure to a stimulus, for example, the differential frequency exposure to kin versus non-kin during a life cycle), trial-and-error learning (i.e. repeated attempts to solve a task until success, for example, attempts to behave altruistically toward kin, and the benefits it entails, versus the costs of maladaptive altruism toward non-kin), or classical or operant conditioning to discriminate between kin and non-kin (i.e. learning to associate a behavioral or chemical cue with the advantages/disadvantages of aggregating, cooperating or reproducing with conspecifics of [diverse degree of relatedness]). All these topics, remain open areas of investigation and experimentation with protists since, like Paramecium, they possess basic sensory perception capabilities, which could have been co-opted [= adapted] during evolution to function in kin discrimination/recognition…”
Social amebas (or facultative social)
The second article (by Strassmann) is a another review, in this case on “Kin Discrimination in Dictyostelium Social Amoebae.” In it, the author recounts her research program on various species of social amebas, including Polysphondylium violaceum, D. purpureum and D. giganteum. Here is a simplified version of the abstract:
“Evolved cooperation is stable only when the benefactor is compensated, either directly or through its relatives. Social amoebae cooperate by forming a mobile multicellular body in which about 20% of participants ultimately dies to form a stalk [watch video below]. This benefits the remaining individuals that become hardy spores at the top of the stalk, together making up [a] fruiting body. In studied species [of social ameba] with stalked migration, P. violaceum, D. purpureum, and D. giganteum, sorting based on clone identity occurs in laboratory mixes, maintaining high relatedness within the fruiting bodies. D. discoideum has unstalked migration where cell fate is not fixed until the slug forms a fruiting body. Laboratory mixes show some degree of both spatial and genotype-based sorting, yet most laboratory fruiting bodies remain chimeric. However, wild fruiting bodies are made up mostly of clonemates. A genetic mechanism for sorting is likely to be cell adhesion genes tgrB1 and tgrC1, which bind to each other. [These genes] are highly variable, as expected for a kin discrimination gene. It is a puzzle that these genes do not cause stronger discrimination between mixed wild clones, but laboratory conditions or strong sorting early in the social stage diminished by later slug fusion could be explanations.”
Above: the amazing videos of social behavior in amebas, by John Bonner, Professor of Biology at Princeton University. He obtained the images as an undergraduate student. This is a 2-min video of historical value, watch it to the end (it turns spectacular).
Schematic phylogeny based on ssrRNA sequences of the Entamoeba clones discussed in the Espinosa et al. article. Free-living: E. moshkovskii Laredo; commensal E. terrapinae and E. dispar; and parasitic E. invadens IP-1, E. invadens VK-1:NS, E. moshkovskii Snake and E. histolytica HM-1:IMSS.
“Studies on clone- and kin-discrimination in protists have proliferated during the past decade. We report clone-recognition experiments in seven Entamoeba [varieties] (E. invadens IP-1, E. invadens VK-1:NS, E. terrapinae, E. moshkovskii Laredo, E. moshkovskii Snake, E. histolytica and E. dispar). First, we characterized morphometrically each clone (length, width, and cell-surface area) and documented how they differed statistically from one another (as per single-variable or canonical-discriminant analyses). Second, we demonstrated that amebas themselves could discriminate self (clone) from different (themselves versus other clones). In mix-cell-line cultures between closely-related (E. invadens IP-1 versus E. invadens VK-1:NS) or distant-phylogenetic clones (E. terrapinae versus E. moshkovskii Laredo), amebas consistently aggregated with same-clone members. Third, we identified six putative cell-signals secreted by the amebas and which known functions in Entamoeba spp. included: cell proliferation, cell adhesion, cell movement, and stress-induced encystation. To our knowledge, this is the first multi-clone characterization of Entamoeba spp. morphometrics, aggregative behavior, and cell-signaling secretion in the context of clone-recognition. Protists allow us to study cell-cell recognition from ecological and evolutionary perspectives. Modern protistan lineages can be central to studies about the origins and evolution of multicellularity.”
Conclusion
Again, I would like to quote the first article: “…because protists are among the most ancient organisms on Earth, belong to multiple taxonomic groups and occupy all environments, they can be central to reexamining traditional hypotheses in the field of kin recognition, reformulating concepts, and generating new knowledge [to our current understanding of the origins and evolution of multicellularity].” – GPC – Evolution Literacy.
You can contact Guillermo Paz-y-Miño-C via email at guillermo.pazyminoc@gmail.com
Protists are among the most ancient organisms on Earth; they belong to multiple taxonomic groups and occupy all environments. Studies with protists can be central to generating new knowledge to our current understanding of the origins and evolution of multicellularity.
* Darwin (1859) speculated about the “puzzle of the sterile social insects,” in which female workers at a nest dedicate their lives to the persistence of the colony (structured around a large progeny), via assisting a fertile queen to reproduce with the available males. Darwin suggested that, in such cases of apparent sacrifice —by the workers— for the good of all, “selection may be applied to the family.” But, in the late 1800s, he could not offer a detailed mechanistic explanation for the latter. Fisher (1930) and Haldane (1932, 1955) wrestled with the genetics and mathematics of altruism and the anecdotic expression “I would lay down my life for two brothers or eight cousins” became legacy of their work. Hamilton (1964) and Maynard-Smith (1964) further reasoned that the ability to discriminate between close and distant genetic relatives could be directly linked to survival and reproductive success, and, ultimately, to kin selection (Maynard-Smith 1964, 1977). — For references, go to source.
The late Intelligent Design, or Design Creationism, continues to decompose. Indeed, pseudo-science has never been able to withstand the power of science. And the story below is quite compelling, not because scientists interested in studying the molecular sophistication of the bacterial flagellum had in mind to further debunk ID. In fact, the researchers (Beeby et al.) did not even mention Design Creationism in their work. But because their study, just published in PNAS, gives US, the people, the opportunity to extrapolate, once more, that there is not a hint of evidence in support to “irreducible complexity” in the anatomy or function of the bacterial flagellum. A reassurance that Design Creationism will forever be wrong. – Guillermo Paz-y-Miño-C
Remember the bacterial flagellum? The pet-example of the late Intelligent Design movement? The alleged “case-study” of irreducible complexity? Well, there are more bad news for Design Creationism. A paper on the bacterial flagellum, recently published in the Proceedings of the National Academy of Sciences (PNAS), explores even further the motor (in reality, the flagellum’s rotor) diversity in two species of bacteria (Campylobacter and Vibrio, plus the study makes comparisons to a third species, Salmonella). Unfortunately, the article is not friendly written, and it can be difficult to understand; the science, however, is exquisite.
Before I summarize the study, watch this 22-sec video about the general structure of the bacterial flagellum. It is simple and will help you understand everything else below:
Here, I try to explain the paper by paraphrasing it: Beeby et al. (a total of six coauthors) have found evidence that “bacteria have tuned their swimming abilities [to their surrounding environments] by evolving structural adaptations to their flagellar motors [which are made of proteins], and that [have resulted] in altered torque generation.” [Note that torque is the twisting force that causes rotation of the flagellum]. In essence, “different bacteria show different swimming [styles], [plus different flagellar-motor anatomies], strikingly illustrated by [their] abilities to bore through viscous fluids (for example, the gastrointestinal mucus) in which other bacteria are [unable to swim].”
There is plenty of evidence of gradually evolved differential complexity among flagellar-motors. Bacterial motors with different torque (Salmonella, Vibrio and Campylobacter). Images by Morgan Beeby Imperial College London (see article in PNAS 2016).
Although the authors say nothing about Intelligent Design in their study, for obvious reasons, i.e. who bothers to allude to a non-scientific proposal (ID) in a serious scientific paper (PNAS). We, the readers, however, can extrapolate from the Beeby et al.’s elegant study that there is NOT a hint of evidence in support to “irreducible complexity” in the form or function of the bacterial flagellum, as proposed by Design Creationists. But rather, there is plenty of evidence of gradually evolved differential complexity among flagellar-motors. This complexity is rooted in ancestry, from absence of the rotor in ancient forms of bacteria, to presence and diversity of rotors in more recent forms of bacteria. That is, “gradual change with modification” (or classical Darwinian evolution), exactly the opposite to the irreducibly-complex flagellum designed by a “Designer of Nature,” the imaginary force of causality invoked by Intelligent Designers. In conclusion, the science story about the bacterial flagellum is, by far, more exciting than the pseudo-science, creationist tale of ID. – Evolution Literacy
The INDEPENDENT “Scientific Paper Sparks Controversy”
Click on image to be redirected to the Independent article.
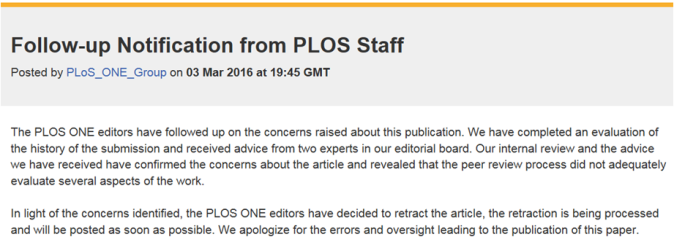
The paper by Liu M-J et al. (Biomechanical Characteristics of Hand Coordination in Grasping Activities of Daily Living) must be retracted from PLoS ONE, and its editor, Renzhi Han (Ohio State University Medical Center), dismissed from the journal for carelessness in processing the manuscript, to say the least. Here is why: the authors (apparent sympathizers of Intelligent Design) invoke the “Creator” in the Abstract, Introduction, Discussion, and also in the Comments section of the online version of their article, in which they respond to criticisms by the readers and editors at PLoS. And this is not an error of translation, nor a struggle in the authors’ attempt to explain, in English, the contrast between the evolutionary origin of the mechanic dexterity of the human hand and its alternative, unsupported hypothesis, “DESIGN CREATIONISM” responsible for it. No, the authors, with the blessings of the journal’s editor, insist that a Creator, or Designer, made the hand almost perfect. This is bad science and terrible editing by PLoS ONE. Below, I summarize the case and provide links to the journal and PDF. But keep in mind that PLoS ONE might retract the article, terminate the editor, and the information from the journal website might be removed. In any event, the paper by Liu M-J et al. must go from PLoS ONE to ZERO. – Guillermo Paz-y-Miño-C
The paper by Liu M-J et al. was received by PLoS ONE on October 28, 2015; accepted on December 14, 2015; and published on January 5, 2016. I missed it, but became aware of it by allusions to it in the social media (Retraction Watch alerted it on March 2, 2016).
Citation: Liu M-J, Xiong C-H, Xiong L, Huang X-L (2016) Biomechanical Characteristics of Hand Coordination in Grasping Activities of Daily Living. PLoS ONE 11(1): e0146193. doi:10.1371/journal.pone.0146193.
What is the paper about?
In the authors’ own words “…This study explores a method to identify the proper explanation for the hand architecture of muscular-articular connections from the analysis of behavioral result…”
Liu M-J et al. examine the biomechanics of the human hand; its dexterity and relationships between anatomy and function. The authors aim at conveying the message that any robotic attempt to mimic the ability of the human hand should pay close attention to its anatomy (and physiology), which, according to Liu M-J et al., would lead to best outcomes in robotic-engineering design. So far, so good. But what follows below is unacceptable, and for that reason the paper must be retracted and its editor, Renzhi Han (Ohio State University Medical Center), dismissed from PLoS ONE.
A hand wearing an instrumented glove demonstrates diverse types of dexterity (click on image to enlarge). Source Liu M-J et al. PLoS ONE 2016.
This is what the authors state in the ABSTRACT:
“Hand coordination can allow humans to have dexterous control with many degrees of freedom to perform various tasks in daily living. An important contributing factor to this important ability is the complex biomechanical architecture of the human hand… It is not understood which biomechanical characteristics are responsible for hand coordination and what specific effect each biomechanical characteristic has. To explore this link, we first inspected the characteristics of hand coordination during daily tasks… from thirty right-handed subjects during a multitude of grasping tasks. Then, the functional link between biomechanical architecture and hand coordination was drawn by establishing the clear corresponding causality between the tendinous connective characteristics of the human hand and the coordinated characteristics during daily grasping activities. The explicit functional link indicates that the biomechanical characteristic of tendinous connective architecture between muscles and articulations is the proper design by the Creator to perform a multitude of daily tasks in a comfortable way. The clear link between the structure and the function of the human hand also suggests that the design of a multifunctional robotic hand should be able to better imitate such basic architecture.”
Thus, Liu M-J et al. invoked “design intervention” to account for complexity.
This is what the authors state in the INTRODUCTION:
“The human hand is an amazing instrument that can perform a multitude of functions, such as the power grasp and precision grasp of a vast array of objects. The excellent behaviors of the human hand are enabled by a highly complex structure, with 19 articulations, 31 muscles and more than 25 degrees of freedom (DOF). While the abundant functions are favorable, this complex structure also raises a challenging problem of how the human body controls such a large number of mechanical DOFs with ease and an absence of effort…”
To help readers understand, we can simply call DOF “dexterity,” or the readiness and grace in physical activity, the skill and ease in using the hands.
Liu M-J et al. continue: “…Studies indicate that digits do not move alone in isolation of adjacent digits during functional activity, even when a specific movement requires an individual digit. On the contrary, the movements of multiple digits are correlated, and movement information of the human hand is redundant, so that only a small number of components account for most variances. The human hand adopts coordinated movements to reduce the number of independent DOFs and simplify the complexity of the control problem. Thus, hand coordination affords humans the ability to flexibly and comfortably control the complex structure to perform numerous tasks. Hand coordination should indicate the mystery of the Creator’s invention…”
Once again, Liu M-J et al. invoked a “Creator’s intervention” to account for complexity.
This is what the authors state in the DISCUSSION:
In closing the article, Liu M-J et al. reaffirm: “…the architecture is the biomechanical basis of the dexterous movement that provides the human hand with the amazing ability to perform a multitude of daily tasks in a comfortable way. In conclusion, our study can improve the understanding of the human hand and confirm that the mechanical architecture is the proper design by the Creator for dexterous performance of numerous functions following the evolutionary remodeling of the ancestral hand for millions of years. Moreover, functional explanations for the mechanical architecture of the muscular-articular connection of the human hand can also aid in developing multifunctional robotic hands by designing them with similar basic architecture.”
Now, this is what Liu M-J et al. and the Academic Editor, Renzhi Han, do not understand about alleged “design” in nature or “design creationism:”
The doctrine of Intelligent Design (ID), or Design Creationism, born in the 1980s, proposes that a Designer is responsible, ultimately, for the assemblage of complexity in biological systems; according to ID, evolution cannot explain holistically the origin of the natural world, nor the emergence of intricate molecular pathways essential to life, nor the immense phylogenetic differentiation of life, and instead ID proposes an intelligent agent as the ultimate cause of nature. In conceptually mistaken, type-I-error-based arguments to discredit evolution, ID has attributed randomness to molecular change, deleterious nature to single-gene mutations, insufficient geological time or population size for molecular improvements to occur, and invoked “design intervention” to account for complexity in molecular structures and biological processes. In 2005, ID was exposed in court (Dover, Pennsylvania, Kitzmiller et al. versus Dover School District et al. 2005) for violating the rules of science by “invoking and permitting supernatural causation” in matters of evolution, and for “failing to gain acceptance in the scientific community” (excerpt from Paz-y-Miño-C and Espinosa 2013, 2014).
And this is what Liu M-J et al. and the Academic Editor at PLoS have done: invoke and permit supernatural causation in matters of evolution.
Notification from PLOS Staff
Yesterday, March 2, 2016, the very journal PLoS ONE posted the following notification on its website (Comments section of article):
“… A number of readers have concerns about sentences in the article that make references to a ‘Creator’. The PLOS ONE editors apologize that this language was not addressed internally or by the Academic Editor during the evaluation of the manuscript. We are looking into the concerns raised about the article with priority and will take steps to correct the published record.”
But the reaction from PLOS just happened, after two months of the paper being on air. Look at this exchange between one of the readers of PLoS ONE and the first author of the paper, Mr. Liu M-J.
“In the abstract (and similarly later in the paper) it is claimed: The explicit functional link indicates that the biomechanical characteristic of tendinous connective architecture between muscles and articulations is the proper design ‘by the Creator’ to perform a multitude of daily tasks in a comfortable way. I am interested to hear from the authors why one should come to this conclusion? I could not find any support for this claim in the article, and it seems out of place in this article…”
“Thanks for your comments. As we know, human hand is an amazing instrument that can perform a multitude of functions, such as the power grasp and precision grasp of a vast array of objects, with ease and an absence of effort. Although expended great attempts by scientists and engineers, there is no artificial hand matching the amazing capacity of human hand. The origins of human hand remain unclear. It is too miraculous to let us think that human hand is the masterwork of Creator and indicates the mystery of nature. The further discussion about the Creator is indeed out of place in our article…”
In addition, today, March 3, 2016, Mr. Liu M-J, posted this note at PLoS Comments:
“We are sorry for drawing the debates about creationism. Our study has no relationship with creationism. English is not our native language. Our understanding of the word Creator was not actually as a native English speaker expected. Now we realized that we had misunderstood the word Creator. What we would like to express is that the biomechanical characteristic of tendious connective architecture between muscles and articulations is a proper design by the NATURE (result of evolution) to perform a multitude of daily grasping tasks. We will change the Creator to nature in the revised manuscript. We apologize for any troubles may have caused by this misunderstanding.”
A bit late, the analogy NATURE = Creator is not new among spiritualists (or even religions), the problem is that it was used in a scientific paper by professionals in a scientific field. And the Academic Editor at PLoS allowed it a complete ride up to publication. And that is the issue we are discussing. And the authors, and editor, must know that a “Creator” (unequivocally invoked three times in the paper and once in the Comments) has no place in Science.
What should PLOS and PLoS ONE do?
It is evident that neither the authors, nor the Academic Editor, understand how evolution via natural selection works. Worse, none is aware of the fallacies intrinsic to Intelligent Design, or Design Creationism. Here a common-knowledge principle applies “not knowing the law does not exempt anyone from having the law being applied to everyone.” What law? At least one and two bylaws: natural selection (which explains the origin and evolution of the human hand), plus the bylaws of publication of articles in scientific journals (i.e. substantiation by evidence, not by the whim of the authors or editors, and sound editorial process). [For accuracy, be aware that natural selection is not a “law,” I am using it here as an unavoidable mechanism that helps us explain much of the evolutionary processes; selection imposes restrictions on randomness and helps us dismiss, quite easily, the fallacy of “design creationism”]. What should be done now that we, the people, caught this “error” at PLoS, after two months since publication?
The allusions to the Creator or Designer by Liu M-J et al. are out of place, but they do invoke and permit supernatural causation in matters of evolution. Unfortunately, there is no evidence of a problem in translation, from the authors’ native language into English. In fact, the authors insist in the Abstract, Introduction, Discussion, and in the Comments section of their article that they do give credit, in a scientific publication, to the long-time debunked hypothesis of Intelligent Design, Design Creationism. The paper must be retracted from PLoS ONE, the Academic Editor, Renzhi Han (Ohio State University Medical Center), must be dismissed from PLOS, and the journal must assure the scientific community that the Public Library of Science (PLoS) will never, ever embrace design creationism in its publications. The paper by Liu M-J et al. must go from PLoS ONE to ZERO. — Evolution Literacy
On March 4, 2016, PLoS ONE officially retracted the paper:
Click on image to be redirected to PLoS ONE retraction announcement.
If you are on Twitter (or Facebook), you have probably asked yourself two questions: Am I less popular than my social-media friends? Am I less happy than them? These phenomena (i.e. cyber popularity or cyber happiness) are both real and perceptual.
A paper, by Bollen et al. (total four coauthors), which is available as manuscript (The Happiness Paradox: Your Friends Are Happier Than You), caught my attention. The authors provide evidence of complex associations between the “friendship paradox” and the “happiness paradox.” Enjoy. – Guillermo Paz-y-Miño-C
What is the Friendship Paradox? The perception that YOU are less popular than your friends (on average).
What is the Happiness Paradox? The perception that YOU are less happy than your friends (on average).
What is the issue (so what)? The friendship paradox has been documented in the scientific literature (see Suggested Readings below), but the happiness paradox has remained untested, without quantitative demonstration that it actually happens. Or that, if it does, it occurs in association –somehow– with the friendship paradox. The paper by Bollen et al. makes such connection.
To make it easier for our readers, I will summarize the Bollen et al. study in a digestible manner, although the authors themselves do a fine job explaining the theory, the data and the implications of the research to a specialized audience (see PDF of manuscript). But, for the non-scientists, my summary below might help.
The observations in nature (the social environment):
Online social networking is associated with elevated levels of loneliness, anxiety, displeasure, and dissatisfaction. For example, many times you are on Twitter (or Facebook, or alike social media), you experience an overall sensation that your averagecontacts (i.e. friends, people you follow, or your followers) have more friends or followers than you do, or are happier than you are.
When measured in number of friends-connections, most people tend to have fewer friends than their own friends do on average. Why? Watch the short video posted below (1 minute 36 seconds of your time) for a friendly introduction to the Friendship Paradox.
Now, let us examine numerically how the Friendship Paradox emerges from a simple example. Imagine a social network of four people, as depicted in the image below: Ana has 1 friend (Bill). Bill has 3 friends (Ana, Carla and Dan). Carla and Dan have 2 friends each (Bill, and each other).
There are 8 friends in the network (1 + 3 + 2 + 2). Thus, on average, each person has 2 friends (8/4 = 2).
However, how many friends does each person’s friends have (i.e. how many friends of friends)? Ana is connected to +3 friends of friends, via Bill (who has 3 friends), a subtotal of 3 friends of friends. Bill is connect to 1 + 2 + 2 friends of friends (Ana has 1, Carla has 2, and Dan has 2), a sub total of 5 friends of friends. Carla is connected to 3 + 2 friends of friends (Bill has 3 and Dan has 2), a sub total of 5 friends of friends. And Dan is connected to 3 + 2 friends of friends (Bill has 3 and Carla has 2), a sub total of 5 friends of friends. Therefore, the grand total of friends of friends in the network is 18.
What is the average number of friends of friends? We obtain this number by dividing the total number of friends of friends in the network (=18) by the total number of friends in the network (=8), which gives us a value of 2.25. The Friendship Paradox becomes evident for Ana, Carla and Dan since their average number of friends in the network (=2.0) is lower than the average number of friends of friends in the network (=2.25). The phenomenon is particularly acute for Ana since she has a single friend (Bill) and, therefore, her friendship relations are clearly below both the average number of friends in the network (2.0) and the average number of friends of friends in the network (2.25).
This overall sampling bias effect (= the Friendship Paradox) results –in part– from Bill’s presence in the network. Bill has the most friends and is more often counted in the friends of friends tabulation, thus rising the average value (to 2.25).
The hypothesis
At least in part, Bollen et al. tested the hypothesis that the friendship and happiness paradoxes are predictable byproducts (effects) of universal social network connectivity patterns. And that associations between friendship and happiness also derived from such connectivity (note that the Friendship Paradox has been demonstrated numerous times, but Bollen et al. demonstrated it again with their sample to be able to link it to the Happiness Paradox, the central aspect of their paper).
The phenomena “friendship paradox” or “happiness paradox” result from the inherent structural bias in social networks that favor popular individuals (like Bill, above), who are, by definition, more likely to belong to someone’s social circle. For example, when individuals like you and I equate popularity of our cyber-friends with their cyber-prestige, and compare our own popularity to theirs, we tend to increase our levels of dissatisfaction with ourselves (i.e. we tend to enhance, in our minds, the effects of the friendship-paradox- or the happiness-paradox).
The predictions
Bollen et al. predicted that the effects of the friendship paradox will extend beyond popularity. If popular individuals tend to be happier, then their elevated happiness will become more prevalent as well. Thus, friendship-levels and happiness-levels (as per both paradoxes) should correlate, and this relationship should be measurable.
The authors also predicted that unhappiness will contribute more directly to the negative psycho-social effects of social networking, since it will affect how individuals assess their own subjective wellbeing (i.e. general happiness/unhappiness or life satisfaction/dissatisfaction relative to that of others).
The friendship paradox or the happiness paradox will generate predictable distributions of data, as shown in Fig. 1, below:
Figure 1. Predictable distributions of data expected if the Friendship or Happiness paradoxes are supported by the data (redrawn and adapted from Bollen et al.).
The study
Bollen et al. examined 40,000 Twitter users connected by reciprocal friendship (i.e. people that follow each other).
Two distinctive groups were characterized, the Unhappy group and the Happy group, by applying a subjective assessment of their wellbeing (derived from the Twitter-users’ interactions with their cyber-neighbors), which ranged in a scale from -1.0 (unhappy) to +1.0 (happy).
As shown in Fig. 2, below, both the Unhappy and Happy groups experienced evident friendship-paradox effects (i.e. members of both groups did have, or thought that their friends had, more friends, on average, than themselves). Interestingly, the intensity of the friendship paradox was weaker in the Unhappy group (left) than in the Happy group (right).
Figure 2. Friendship-paradox effects on the Unhappy (left) and Happy (right) groups. Note how the intensity of the friendship paradox was weaker in the Unhappy group than in the Happy group. The “clouds” of data correspond to point distributions of each of the 40,000 Twitter users in the study (modified from Bollen et al.).
In addition, as Fig. 3, below, suggests, both the Happy (top) and Unhappy (bottom) groups experienced clear happy-paradox effects. However, the intensity of the happiness paradox was stronger in the Unhappy group (see the very steep slope in the data point distribution, plus the amount of data points over the main line) than in the Happy group (see the not-so-steep slope in the data point distribution, plus the amount of data points over the main line). In essence, the majority of the sampled Twitter users were less happy (or more unhappy) than their friends on average. These findings support the idea that the Happiness Paradox is real.
Figure 3. Scatter point distributions of an individual’s (exemplified by YOU) sense of happiness as function of his/her (exemplified by YOUR) friends’ average sense of happiness (modified from Bollen et al.).
Individual Happiness and average friends’ Happiness were more strongly related within the Unhappy group than within the Happy group (Fig. 3). Bollen et al. refer to the clustering of unhappy users with themselves, and happy users also with themselves, as being homophilic or assortative in their social networking (both are common terms in behavioral sciences).
Figure 4, below, corroborates the finding that Unhappy Twitter users had fewer friends, on average, than the Happy Twitter users had, on average.
Figure 4. Unhappy Twitter users had fewer friends, on average, than the Happy Twitter users had, on average (modified from Bollen et al.).
Figure 5, below, corroborates the finding that the Happiness Paradox affected more the Unhappy Twitter users, on average, than the Happy Twitter users, on average.
Figure 5. The Happiness Paradox affected more the Unhappy Twitter users, on average, than the Happy Twitter users, on average (modified from Bollen et al.).
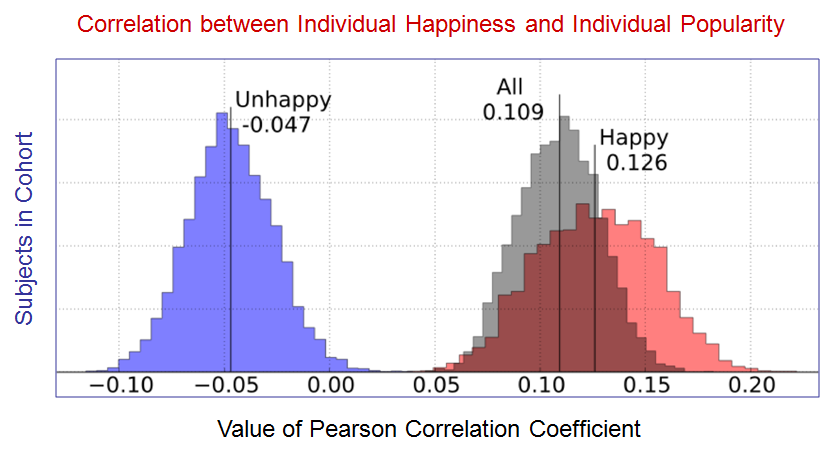
Paradoxically, however, as shown in Fig. 6, below, there was a weak correlation between individual happiness and individual popularity for the Unhappy group (Pearson’s R = -0.047; range -0.08 to -0.013). For the Happy group, the correlation was more pronounced (Pearson’s R = 0.126; range 0.081 to 0.171).
Figure 6. There was a weak correlation between individual happiness and individual popularity for the Unhappy group. For the Happy group, the correlation was more pronounced (modified from Bollen et al.).
The overall results suggest that unhappy Twitter users were more strongly affected by the lower happiness of their friends, possibly explaining why this group exhibited a stronger Happiness Paradox (Figs. 3 and 5) in the absence of a strong correlation between Happiness and Popularity (Fig. 6).
Observations of decreased happiness among social media users may result directly from a widespread inflated perception of the happiness of one’s friends.
Given the large role that social media (e.g. Twitter, Facebook) plays in the social lives of billions of individuals, the authors suggest that these environments may induce long-term changes in the public’s social behavior and may, over time, alter the very nature of social relations themselves. — EvoLiteracy.
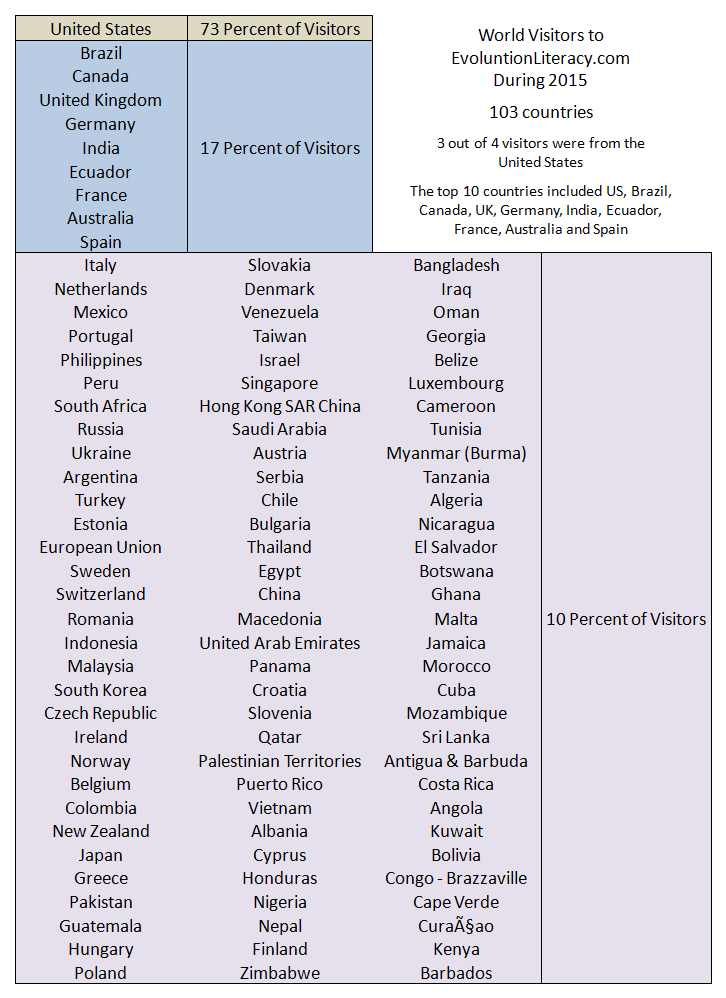
World Visitors to EVOLUTION LITERACY – Readers from 103 countries visited EvoLiteracy during 2015. Three in every four readers were from the United States. About one in every five visitors were from Brazil, Canada, UK, Germany, India, Ecuador, France, Australia and Spain. And one in every ten cyberworms came from 93 other countries. EvoLiteracy is growing thanks to our world friends and followers. – Guillermo Paz-y-Miño-C.
There are 190+ countries in the world (member states of the United Nations). EvoLiteracy reaches half of them (52%). The image below includes flags of nations, which total 230+. We still need to reach as many nations as possible. Please share EvoLiteracy with others.
Flags of 230+ nations in the world. Click on image to enlarge. Source Danilka’s Blog.
Top 25 Most Read Posts of 2015
Here are the most popular postings of 2015. I was glad to discover that the biology science videosmade much of an impact, particularly among science educators. I also liked that three crucial postings about higher education (marked with an asterisk * below) were well received. To my surprise (and I thank the readers for liking it), the posting about Ecuador’s Academy of Science was ranked top 10. My personal favorite was Science Challenges Golden Age of Violin Making, and this is because I am fascinated with string instruments (classic guitars, ukuleles, charangos); I learned much while investigating the violin ancestry. But I cannot close without admitting how much pleasure gave me to see our readers liking Evolution Wars Debunk II (ranked 13th, a lucky number). Plus the most commented story was Shroud of Turin, Poor Science, and the Persistence of a Myth, which was reposted in various blogs and generated two weeks of discussions. Thanks to all for supporting EvoLiteracy. – GPC
14 EvoLiteracy News 05 08 2015 Should scientific journals request authors to change their practices for presenting continuous data in small sample size studies?
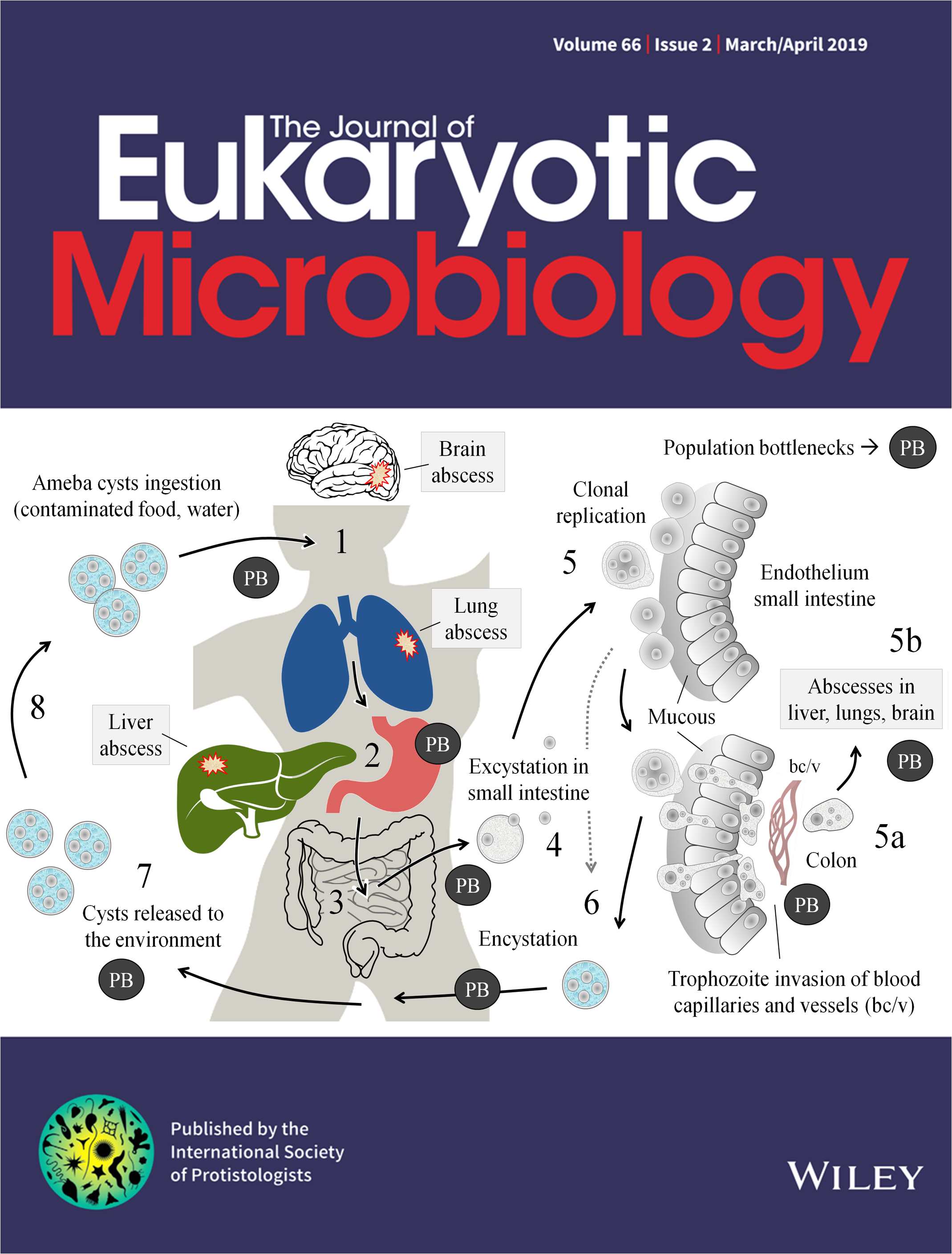 Our latest review on clone-clone discrimination/recognition in Entamoeba species has been featured on the cover of the Journal of Eukaryotic Microbiology (JEUK-MIC, Volume 66, Issue 2, March-April 2019). This is the second time our work has made it to the cover of JEUK-MIC (the first time was back in 2012, coincidentally in the March-April Volume 59, Issue 2). Below are the links to the 2019 article, as well as the abstract and caption to the cover image.
Our latest review on clone-clone discrimination/recognition in Entamoeba species has been featured on the cover of the Journal of Eukaryotic Microbiology (JEUK-MIC, Volume 66, Issue 2, March-April 2019). This is the second time our work has made it to the cover of JEUK-MIC (the first time was back in 2012, coincidentally in the March-April Volume 59, Issue 2). Below are the links to the 2019 article, as well as the abstract and caption to the cover image.

































You must be logged in to post a comment.